Считается, что фотосинтез развился несколько миллиардов лет назад из аноксического фотосинтетического комплекса у предка, давшего начало современным цианобактериям.
Тилакоиды: характеристики, строение и функции

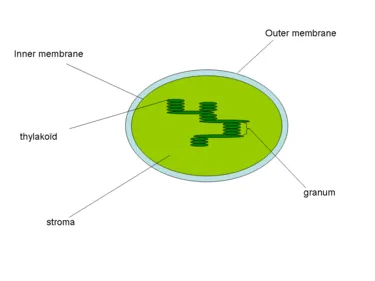
Тилакоиды — это плоские, похожие на мешочки камеры, которые находятся в хлоропластах клеток растений, цианобактерий и водорослей. Они обычно организованы в структуру под названием granum — множественное число granum — которая напоминает стопку монет.
Тилакоиды считаются третьей мембранной системой хлоропластов, наряду с внутренней и внешней мембранами этой органеллы. Мембрана этой структуры отделяет внутренний тилакоид от слоя хлоропласта и содержит различные пигменты и белки, участвующие в метаболических процессах.
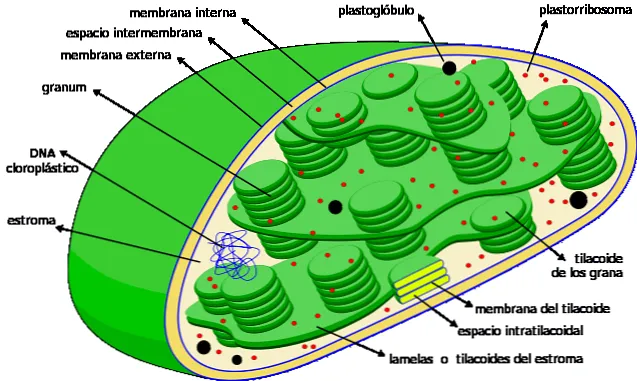
В тилакоидах происходят биохимические реакции, необходимые для фотосинтеза — процесса, в ходе которого растения поглощают солнечный свет и преобразуют его в углеводы. В частности, они имеют на своей мембране необходимое оборудование для осуществления зависимой фазы захвата солнечного света и преобразования его в энергию (АТФ) и NADPH.
Общие характеристики
Тилакоиды — это внутренняя трехмерная мембранная система хлоропластов. Полностью зрелые хлоропласты содержат от 40 до 60 гранул в стопке диаметром от 0,3 до 0,6 мкм.
Количество тилакоидов, входящих в состав растений, сильно варьирует: от менее 10 тилакоидов у растений, подвергающихся достаточному воздействию солнечного света, до более 100 тилакоидов у растений, живущих в чрезвычайно затененной среде.
Уложенные друг на друга тилакоиды соединены между собой и образуют целостный отсек внутри хлоропласта. Внутренняя часть тилакоида представляет собой довольно просторный водный отсек.
Тилакоидная мембрана необходима для фотосинтеза, так как здесь происходит первый этап процесса.
Состав
Тилакоиды являются доминирующими структурами в полностью зрелом хлоропласте. При визуализации хлоропласта под обычным световым микроскопом можно выделить несколько типов зерен.
Это стопки тилакоидов, поэтому ранние наблюдатели этих структур называли их «грана».
С помощью электронного микроскопа изображение можно было увеличить, и был сделан вывод, что эти зерна действительно являются естественно сложенными тилакоидами.
Образование и структура тилакоидной мембраны зависит от формирования хлоропласта другой недифференцированной пластидой, называемой протоплазмидой. Присутствие света стимулирует превращение в хлоропласты и последующее образование фолликулов.
Общие характеристики
Тилакоиды — это внутренняя трехмерная мембранная система хлоропластов. Полностью зрелые хлоропласты имеют от 40 до 60 гранул диаметром от 0,3 до 0,6 мкм.
Количество тилакоидов, из которых состоят гранулы, сильно варьирует: от менее 10 мешочков у растений, подвергающихся достаточному воздействию солнечного света, до более чем 100 тилакоидов у растений, живущих в условиях экстремальной тени.
Уложенные друг на друга тилакоиды соединены между собой и образуют целостный отсек внутри хлоропласта. Внутренняя часть тилакоида представляет собой довольно просторный водный отсек.
Тилакоидная мембрана необходима для фотосинтеза, поскольку именно в ней происходит первый этап…
Структура
Тилакоиды являются доминирующими структурами в полностью зрелом хлоропласте. Если вы изобразите хлоропласт с помощью обычного светового микроскопа, вы сможете увидеть определенные типы гранул.
Это стопки тилакоидов, поэтому ранние наблюдатели этих структур называли их «грана».
С помощью электронного микроскопа они смогли увеличить изображение и пришли к выводу, что эти зерна на самом деле являются свернутыми тилакоидами.
Образование и структура тилакоидной мембраны зависит от формирования хлоропласта из еще недифференцированной пластиды, так называемой протоплазмиды. Присутствие света стимулирует превращение в хлоропласты и последующее образование фолликулов.
Тилакоидная мембрана
В хлоропластах и цианобактериях тилакоидная мембрана не имеет контакта с внутренней частью плазматической мембраны. Однако формирование мембраны фолликула начинается с проникновения внутренней мембраны.
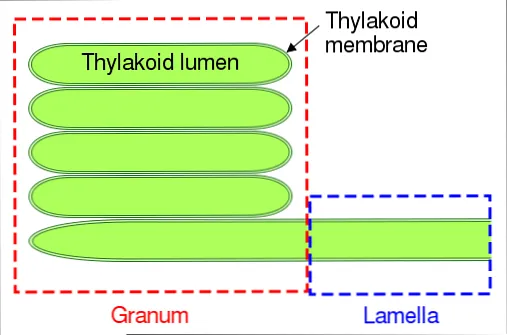
У цианобактерий и некоторых видов водорослей фолликулы состоят из одного слоя ламелей. В зрелых хлоропластах, с другой стороны, система более сложная.
В последней группе можно выделить две основные части: зернистый слой и пластинчатый слой. Первый состоит из маленьких дисков, уложенных друг на друга, а второй отвечает за соединение этих слоев вместе, образуя непрерывную структуру — фолликулярный просвет.
Липидный состав мембраны
Липиды, составляющие мембрану, высокоспециализированы и почти на 80% состоят из галактозилдиацилглицерина: моногалактозилдиацилглицерина и дигалактозилдиацилглицерина. Эти галактолипиды имеют высоко ненасыщенные цепи, характерные для тилакоидов.
Аналогичным образом, тилакоидная мембрана содержит липиды, такие как фосфатидилглицерин, в меньшей пропорции. Эти липиды неравномерно распределены в обоих слоях мембраны; существует некоторая асимметрия, которая, по-видимому, вносит вклад в функционирование структуры.
Белковый состав мембраны
Фотосистемы I и II являются преобладающими белковыми компонентами этой мембраны. Они связаны с цитохромом b6F и АТФ-синтазного комплекса.
Было обнаружено, что большинство элементов фотосистемы II находятся в сложенных мембранах гранул, в то время как фотосистема I в основном находится в неуложенных мембранах тилакоидов. Это означает, что между двумя фотосистемами существует физическое разделение.
Эти комплексы включают целые мембранные белки, периферийные белки, кофакторы и различные пигменты….
Люмен тилакоида
Внутренняя часть тилакоида состоит из водного и плотного вещества, которое отличается по составу от стромы. Он участвует в фотофосфорилировании, накапливая протоны, которые создают протонную движущую силу для синтеза АТФ. Во время этого процесса pH просвета может достигать 4.
В протеоме просвета модельного организма Arabidopsis thaliana идентифицировано более 80 белков, но их функции еще не до конца понятны.
Белки просвета вовлечены в регуляцию биогенеза тилакоидов, а также в активность и метаболизм белков, образующих фотосинтетические комплексы, особенно фотосистемы II и NAD(P)H дегидрогеназы.
Функции
Процесс фотосинтеза, который жизненно важен для овощей, начинается в тилакоидах. Мембрана, окружающая их в слое хлоропласта, содержит все ферментативные механизмы, необходимые для осуществления фотосинтетических реакций.
Этапы фотосинтеза
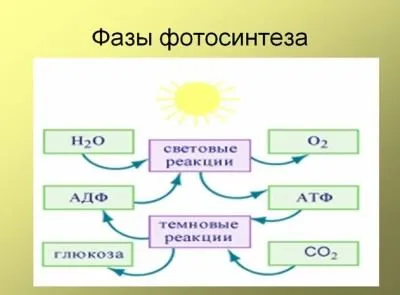
Фотосинтез можно разделить на две основные фазы: Световые реакции и темные реакции.
Как следует из названия, реакции первой группы могут протекать только в присутствии света, в то время как реакции второй группы могут протекать как со светом, так и без него. Не обязательно, чтобы среда была «темной»; она зависит не только от света.
Первая группа реакций, «фосфоресцирующие» реакции, происходят в тилакоиде и могут быть обобщены следующим образом: Свет + хлорофилл + 12 H2O + 12 NADP + + 18 ADP + 18 PЯa 6 o2+ 12 NADP + 18 ATP.
Вторая группа реакций происходит в слое хлоропластов и использует АТФ и NADPH, синтезированные на первом этапе, для преобразования углерода из углекислого газа в глюкозу (C6H12О6Второй этап можно представить следующим образом: 12 NADPHN + 18 ATF + 6 CO2à C6H12О6+ 12 NADP + + 18 ADP + 18 PЯ+ 6 ч2О.
Этап зависит от света
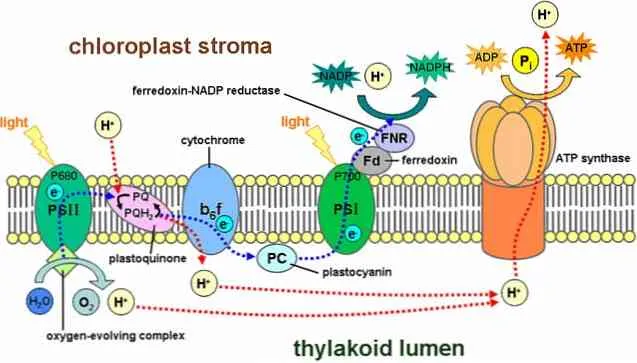
В световых реакциях участвует ряд структур, называемых фотосистемами. Они расположены в тилакоидной мембране и содержат около 300 пигментных молекул, включая хлорофилл.
Существует два типа фотосистем: Первый имеет максимальный пик поглощения света при 700 нанометрах и известен как P700, в то время как второй называется P680. Оба они встроены в тилакоидную мембрану.
Процесс начинается, когда один из пигментов поглощает фотон, и он «отражается» от других пигментов. Когда молекула хлорофилла поглощает свет, электрон отскакивает от нее и поглощается другой молекулой. Молекула, потерявшая электрон, теперь окислена и имеет отрицательный заряд.
P680Он улавливает световую энергию от хлорофилла a. В этой фотосистеме электрон перебрасывается в более высокую энергетическую систему в первичном акцепторе электронов.
Этот электрон попадает в фотосистему I через цепь переноса электронов. Эта система реакций окисления и восстановления отвечает за перенос протонов и электронов от одной молекулы к другой.
Другими словами, существует поток электронов от воды к фотосистеме II, к фотосистеме I и к NADPH.
фотофосфорилирование
Часть протонов, образующихся в этой реакционной системе, находится внутри тилакоидов (также называемых тилакоидным светом), создавая химический градиент, который является движущей силой для протонов.
Протоны перемещаются из фолликулярного пространства в слой, преимущественно следуя электрохимическому градиенту, т.е. покидают фолликул.
Однако прохождение протонов происходит не где-нибудь в мембране, а должно пройти через сложную ферментативную систему, называемую АТФ-синтазой.
Это перемещение протонов в строму запускает образование АТФ, которое начинается с АДФ, процесс, аналогичный тому, что происходит в митохондриях. Синтез АТФ с помощью света называется фотофосфорилированием.
Эти этапы происходят одновременно: Хлорофилл в фотосистеме II теряет электрон и должен заменить его электроном, полученным в результате распада молекулы воды; фотосистема I удерживает свет, окисляет его и отдает электрон, который захватывается NADPH+.
Образование тилакоидов
Хлоропласты развиваются из пропластид, когда побег поднимается над поверхностью почвы. Свет необходим для формирования тилакоидов. В растительном эмбрионе, а также в отсутствие света пропластиды трансформируются в этиопласты, которые имеют полукристаллические мембраны, называемые проламеллярными телами. Под воздействием света эти проламеллярные тела превращаются в тилакоиды. Однако это не относится к побегам, прорастающим в темноте; эти побеги испытывают причинность. Недостаток света может привести к нарушению формирования тилакоидов. Это приводит к разрушению хлоропластов и, как следствие, к гибели растения.
Белок VIPP1 (vesicle-inducing protein in plastids 1) также необходим для формирования тилакоидов. Без этого белка растение погибает, а снижение его концентрации замедляет рост растения, приводит к бледному цвету растения и снижению фотосинтетической активности. VIPP1 необходим для формирования самой тилакоидной мембраны, но не для сборки белковых комплексов в этой мембране 6 .
Этот белок высоко консервативен во всех тилакоид-содержащих организмах, даже в сине-зеленых водорослях, 7 зеленых водорослях, таких как хламидомонады 8 и высших растениях, таких как арабидопсис 9 .
Выделение и фракционирование тилакоидов
Тилакоиды могут быть выделены из растительной клетки с помощью комбинации дифференциального и градиентного центрифугирования 10. Разбив изолированные тилакоиды, например, с помощью механического воздействия, можно выделить вещество просвета. Поверхностные и интегральные белки также могут быть извлечены из оставшихся фрагментов мембраны: Обработка карбонатом натрия (Na2CO3обработка удаляет поверхностные мембранные белки, в то время как обработка моющими средствами и органическими растворителями позволяет извлечь интегральные мембранные белки.
Тилакоиды содержат множество интегральных и поверхностных мембранных белков. Многие белки также присутствуют в просветном веществе. Недавние протеомные исследования 11 фракций тилакоидов дали много информации о белковом составе тилакоидов. Эти данные обобщены в различных базах данных по пластидным белкам, доступных в Интернете 12 13 .
Протеом тилакоидов состоит по меньшей мере из 335 различных белков, как показали исследования, упомянутые выше. Из них 89 находятся в просветном веществе, 116 — интегральные мембранные белки, 68 — на поверхности внутренней мембраны (со стороны просвета) и 62 — поверхностные белки наружной мембраны (со стороны хлоропластного слоя). Кроме того, другие редкие белки просвета были предсказаны с помощью вычислительных методов 14 10. Из тилакоидных белков с известными функциями 42 % участвуют в фотосинтезе. 11 % участвуют в транспорте, обработке и поддержке сворачивания белка, 9 % — в ответе на окислительный стресс и 8 % — в трансляции 12 .
Интегральные мембранные белки
Мембраны тилакоидов содержат интегральные белки, которые играют важную роль в захвате фотонов и светозависимых реакциях фотосинтеза. В мембране есть четыре основных белковых комплекса:
- фотосистемы I и II
- Цитохромный комплекс b6f
- АТФ-синтаза
Фотосистема II находится в основном в гранулярных тилакоидах, а фотосистема I и АТФ-синтаза — в стромальных тилакоидах и во внешних слоях тел гранул. Комплекс цитохрома b6f равномерно распределен в мембране.
Поскольку две фотосистемы пространственно разделены в фолликулярной мембране, для обмена электронами между ними необходимы кинетические транспортеры. Пластохинон и пластоцианин являются такими носителями. Пластохинон передает электроны от фотосистемы II к комплексу b6f-цитохрома, а пластоцианин передает их от комплекса b6f-цитохрома к фотосистеме I.
Вместе эти белки преобразуют световую энергию для приведения в действие цепей электропереноса, которые создают электрохимический потенциал в тилакоидной мембране и синтезируют никотинамид аденоаденин динуклеотид фосфат (NADPH), продукт конечной окислительно-восстановительной реакции. АТФ-синтаза использует этот электрохимический потенциал для синтеза АТФ в процессе фотофосфорилирования.
Фотосистемы
Фотосистемы тилакоидов являются центрами светозависимых окислительно-восстановительных реакций. Каждая фотосистема содержит антенный комплекс, который улавливает свет различных длин волн с помощью хлорофилла и вспомогательных фотосинтетических пигментов, таких как каротиноиды и фикобилипротеины. Антенный комплекс состоит из 250-400 пигментных молекул. Поглощенная ими энергия передается путем резонансной передачи специализированному хлорофиллу a, который находится в реакционном центре каждой фотосистемы. Когда одна из двух молекул хлорофилла a в реакционном центре получает энергию, электрон передается акцепторной молекуле.
Функции тилакоидов
Светозависимые реакции фотосинтеза в тилакоидной мембране
В тилакоидах осуществляются следующие фотозависимые реакции фотосинтеза:
- Светозависимое расщепление воды, в результате которого происходит синтез молекул кислорода;
- Перенос протонов через тилакоидную мембрану, связанный с электронтранспортной цепью фотосистем и цитохромного комплекса b6f;
- Синтез АТФ, выполняемый АТФ-синтазой с использованием протонного градиента.
Фотолиз воды
Первым шагом фотосинтеза является расщепление воды под действием света. Эта реакция обеспечивает электроны для фотосинтетических электронно-транспортных цепей, а также протоны для создания протонного градиента. Реакция расщепления воды происходит на стороне тилакоидной мембраны, обращенной к просвету, и осуществляется за счет потребления энергии, которую фотосинтетическая система получает от солнечного света. Интересно отметить, что это окисление (разложение) воды сопровождается выделением O2в качестве побочного продукта, который выбрасывается в атмосферу и затем может быть использован другими организмами для дыхания.
Электронтранспортная цепь
Основная статья: Электронно-транспортная цепь фотосинтеза
В фотосинтезе используются два типа транспорта электронов:
- Нециклический электронный транспорт или нециклическое фотофосфорилирование, при котором производится NADPH, протоны H + и АТФ;
- Циклический электронный транспорт или циклическое фотофосфорилирование, при котором производится только АТФ.
В некруговом транспорте участвуют обе фотосистемы, тогда как в круговом — только фотосистема I.
- Фотосистема I использует энергию света для восстановления NADP + до NADPH + H +. Она активна в обеих разновидностях электронного транспорта. В циклическом режиме возбужденный электрон отправляется по цепочке, которая в конце возвращает его обратно к хлорофиллу, который сообщил ему энергию возбуждения.
- Фотосистема II использует энергию света для расщепления молекул воды с выходом электронов (e − ), протонов (H + ) и молекулярного кислорода (O2Он участвует только в некруговом транспорте. В этой системе электроны берутся из окисленного 2H2O (O2+ 4 H + + 4 e − ) и удаляются из нее с NADP +, когда он восстанавливается до NADPH.
Электрохимический потенциал
Основная функция тилакоидной мембраны и интегрированных в нее фотосистем заключается в создании электрохимического потенциала. Электронные транспортеры, участвующие в транспорте электронов, используют часть энергии электронов для перекачки протонов из слоя в просвет тилакоида. Во время фотосинтеза просвет закисляется до pH 4 (в то время как строма имеет pH 8). Это соответствует 10 000-кратному градиенту концентрации протонов через тилакоидную мембрану.
Источник протонного градиента
Протоны поступают в просвет из трех источников:
- Фотолиз воды, осуществляемый фотосистемой II, в процессе которого вода в люмене тилакоида окисляется с выходом молекулярного кислорода, протонов и электронов.
- Передача электрона от фотосистемы II к пластохинону во время светозависимой реакции фотосинтеза, входящей в нециклическую электронтранспортную цепь, потребляет два протона из стромы. Они переносятся в просвет тилакоида, когда восстановленный пластохинон окисляется цитохромным комплексом b6f на стороне тилакоидной мембраны, обращенной к просвету. От пластохинона электроны передаются цитохромному комплексу b6f, который напоминает цитохром bc1.
- Восстановление пластохинона ферредоксином в процессе циклического электронного транспорта также вызывает перенос двух протонов из стромы в просвет тилакоида.
Тилакоиды в водорослях и цианобактериях
Хотя в растительных клетках тилакоиды образуют сложенные гранулы, в некоторых видах водорослей они могут разрушаться.
В то время как водоросли и растения являются эукариотами, цианобактерии относятся к фотосинтезирующим прокариотам. Они не содержат хлоропластов. Вместо этого вся клетка функционирует как своего рода тилакоид. Цианобактерии имеют внешнюю клеточную стенку, клеточную мембрану и тилакоидную мембрану. Внутри этой мембраны находятся бактериальная ДНК, цитоплазма и карбоксисомы. В тилакоидной мембране находятся функциональные электронно-транспортные цепи, которые поддерживают фотосинтез и клеточное дыхание. Тилакоидные мембраны цианобактерий не образуют гранул или листа. Вместо этого мембрана образует параллельные листы, прилегающие к цитоплазматической мембране, с достаточным пространством между каждым листом для фикобилисом — структур, собирающих свет.
Где происходит «рождение» энергии и кислорода
Тилакоидная мембрана — это технологическая установка, в которой происходит фотосинтез. Эта реакция происходит через пигмент. Зеленый пигмент хлорофилла поглощает в основном синий и немного красного света из солнечного спектра, что окрашивает отраженный свет в зеленый цвет, т.е. получается тот самый зеленый цвет, который мы видим.
Как и митохондрия, хлоропласт также прокачивает протоны через мембрану. Но в то время как митохондрии транспортируют протоны наружу, протоны собираются в пластидах внутри гранул. Таким образом, мембраны гранул напоминают мембраны митохондрий «вверх ногами». Оба выполняют одну и ту же функцию — преобразование одной энергии (света) в другую — энергию для синтеза АТФ (кислоты, которая обеспечивает энергией химические реакции в клетке).
Отсюда и функции мембран:
- Реакция фотосинтеза. Светозависимое расщепление воды с выделением кислорода.
- Перенос протонов – транспортировка энергии.
- Синтез АТФ (аденозинтрифосфат) – универсального источника энергии для биохимических процессов. Этот процесс очень похож на синтез АТФ в митохондрии.
Бактерии, способные производить кислород
Тилакоиды (ламеллы) встречаются не только у растений, но и у некоторых микроорганизмов — цианобактерий. У цианобактерий нет клеточного ядра и отдельных клеточных структур (органелл), то есть они являются прокариотическими бактериями. Однако цианобактерии содержат структуры, которые позволяют им осуществлять процесс фотосинтеза.

Ученые считают, что хлоропласты произошли от цианобактерий. Это подтверждается тем, что они имеют две мембраны, ДНК и РНК, систему производства белка (хотя и под контролем ядра) и размножаются делением, а их тилакоиды напоминают соответствующие органеллы (структуры) цианобактерий.
Таким образом, эти бактерии похожи на независимые хлоропласты. В отличие от пластид растений, бактериальные ламеллы не объединены по граням, но это не мешает им выполнять свои функции.
Цианобактерии считаются самыми древними бактериями на нашей планете. Их способность утилизировать углекислый газ и производить кислород — это то, что сделало жизнь на Земле возможной в первую очередь.
Три миллиарда лет назад, как сообщают ученые, на нашей планете произошло революционное событие, когда квант солнечного света заставил бактерию расщепить молекулу воды на водород и кислород. С тех пор маленькая желтая звезда, Солнце, стала основным источником энергии для жизни на Земле.
Высшее образование в области литературы. Пишу с 2012 года, также занимаюсь редактированием/публикацией статей. Мои хобби — психология и кулинария.







