Добавление H1, в свою очередь, сворачивает бисерную структуру в нити в спиральную структуру диаметром 30 нм, которая называется 30 нм волокном или нитью. Точная структура хроматиновой нити в клетке детально не известна и до сих пор вызывает споры.
Хроматин это в биологии
Под световым микроскопом мембрана, окружающая ядро, выглядит как единая мембрана, но на самом деле это ядерная оболочка, состоящая из двух мембран. Наружная мембрана заканчивается непосредственно эндоплазматическим ретикулумом (ЭР), как показано на рисунке, и, как и ЭР, может перемежаться с рибосомами, где происходит синтез белка. Ядерная мембрана заполнена ядерными порами. Они особенно заметны в препаратах, изготовленных методом сублимационной сушки. Через ядерные поры происходит обмен различными веществами между ядром и цитоплазмой, например, матричной РНК (мРНК) и рибосомальными субчастицами из цитоплазмы или рибосомальными белками, нуклеотидами и молекулами, регулирующими активность ДНК (например, некоторыми гормонами), в ядро. Поры имеют специфическую структуру, возникающую в результате слияния внешней и внутренней мембран ядерной оболочки. Эта структура регулирует прохождение молекул через поры.
Хроматин
Хроматин состоит из множества цепочек ДНК, соединенных с гистонами, основными белками. Цепочки ДНК настолько длинны (в каждом ядре клетки человеческого тела средняя длина составляет около 1 метра!), что они должны быть уложены упорядоченно, иначе они спутываются, как размотанный клубок. Гистоны и ДНК объединяются и образуют структуры, похожие на бусины, называемые нуклеосомами. Расположение нуклеосом в хроматине также является регулярным.

Слово «хроматин» означает «окрашенный материал» и названо так потому, что хроматин слегка окрашен, когда его готовят для изучения под световым микроскопом. Когда ядро делится, хроматин становится более окрашенным и, следовательно, более видимым за счет конденсации, т.е. образования более плотно закрученных (спиральных) нитей — хромосом. Во время интерфазы (период между двумя делениями ядра) хроматин переходит в более распределенное состояние. Однако сегмент остается плотно закрученным и по-прежнему сильно пигментированным. Этот сегмент называется гетерохроматином; гетерохроматин имеет форму характерных темных пятен, обычно расположенных ближе к ядерной мембране. Остальной, более рыхлый спиралевидный хроматин называется эухроматином.
Считается, что она содержит ДНК, которая генетически активна в интерфазе.
Ядрышко
Ядрышко — это хорошо заметная круговая структура внутри ядра, в которой формируются рибосомы. Ядро может состоять из одного или нескольких ядрышек. Ядро интенсивно окрашено, поскольку содержит большое количество ДНК и РНК. РНК очень похожа на структуру ДНК, потому что она «транскрибируется» вместе с ДНК. Ядро клетки имеет особенно плотную область, где находится ДНК одной или нескольких хромосом. Здесь сосредоточено множество копий генов, кодирующих рибосомальную РНК (рРНК). Во время деления ядро становится невидимым, так как ДНК распределяется. В конце деления ядро вновь появляется.
Центральная область ядра окружена менее плотной периферической областью, где рибосомальная РНК начинает свертываться и происходит сборка рибосомы — РНК связывается с белком. Неполностью собранные рибосомы проходят через ядерные поры из ядра в цитоплазму, где их сборка завершается.
Информация на этом сайте подлежит обсуждению с врачом и не является заменой личной консультации с врачом. Более подробная информация содержится в пользовательском соглашении.
Основные свойства хроматина
Диапазон свойств зависит от типа вещества. Классификация хроматина включает 2 типа. К ним относятся:
- Эухроматин, главной характеристикой которого выступает низкая плотность.
- Гетерохроматин, особенностью которого является повышенная плотность.
Эуфроматин воздействует на ДНК, обеспечивает репликацию — образование двух дочерних молекул ДНК из родительской — и транскрипцию — перенос генетической информации с ДНК на РНК. Эти процессы важны для синтеза ДНК и белков в клетке и для формирования органелл.
Эухроматин обладает рядом свойств, которые позволяют ему осуществлять большинство генетических процессов. Среди них:
- способность постоянно находиться в конденсированном состоянии;
- высокая плотность расположения генов;
- множество копий небольших нетранскрибируемых участков ДНК.
Гетерохроматин, который характеризуется высокой плотностью по сравнению с эухроматином, не способен осуществлять многие генетические процессы. Более того, он присутствует в ядрах женских клеток, в то время как в ядрах мужских клеток он отсутствует. Поэтому многие биологи считают, что это вещество не является генетически активным.
Строение хроматина, химический состав
Структура хроматина меняется в зависимости от периода и фазы клеточного цикла. На уровне промежуточной фазы это глобулы, расположенные по принципу случайной локализации в нуклеоплазме клеточного ядра. При переходе клетки к митозу вещество принимает форму хромосом — отдельных, ярко окрашенных тел.
Основной структурной единицей вещества является нуклеосома, которая состоит из ядрышка, сформированного из двух типов гистонов, вокруг которого обернуты 166 пар оснований дезоксирибонуклеиновой кислоты.
Биохимически состав вещества на 40% состоит из дезоксирибонуклеиновой кислоты и на 60% из белков. Общее количество белков составляет 33% негистоновых белков и 67% гистонов, которые включают H4, H3, H2b, H2a и H1.
Гистоны — это тип белка, который может связываться с ДНК и участвует в ее конденсации и сворачивании.
Стадии конденсации хроматина
Диаграмма конденсации хроматина выглядит следующим образом:
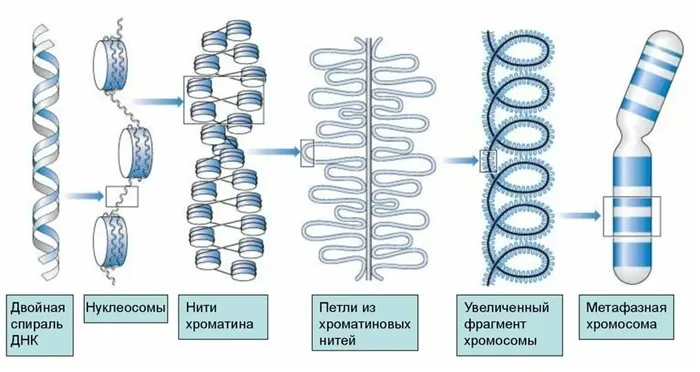
Разные полосы ДНК могут иметь разную степень упаковки в одно и то же время, но каждая полоса неизбежно проходит через 5 стадий:
- Нуклеосомный, в ходе которого ДНК распадается на повторяющиеся структуры из 8 молекул гистонов, известные под названием «октамеры», и намотанную на каждую из них ДНК. Полученные частицы называются нуклеосомами, представляют собой белковую основу с двумя витками ДНК на поверхности.
- Нуклеомерный, завершающийся образованием фибрилл – нитей.
- Хромомерный, на котором формируются петлевые домены ДНК.
- Хроматидный, обеспечивающий дальнейшую компактизацию.
- Уровень метафазной хромосомы.
На последнем этапе хроматиды завершают спирометрию, в результате чего цепь становится в 10 000 раз короче своего первоначального значения.
Конденсация хроматина считается одной из форм апоптоза.
Хроматин в митозе:
Профаза
В процессе митоза хроматиновые волокна превращаются в хромосомы. Каждая реплицирующаяся хромосома состоит из двух хроматид, соединенных с центромерой.
Метафаза
Во время метафазы хроматин сильно сжимается. Хромосомы выравниваются на метафазной пластинке.
Анафаза
Во время анафазы парные хромосомы (сестринские хроматиды) разделяются и притягиваются к противоположным полюсам клетки микротрубочками веретена расщепления.
Телофаза
Во время эндофагии каждая новая дочерняя хромосома мигрирует в свое собственное ядро. Волокна хроматина разматываются и становятся менее толстыми. После цитокинеза образуются две генетически идентичные дочерние клетки. Каждая клетка имеет одинаковое количество хромосом. Хромосомы продолжают раскручиваться и удлинять образовавшийся хроматин.
Хроматин, хромосома и хроматида
Люди часто испытывают трудности с различением терминов «хроматин», «хромосома» и «хроматида». Хотя все три структуры состоят из ДНК и расположены в ядре клетки, каждая из них определяется отдельно.
Хроматин состоит из ДНК и гистонов, упакованных в тонкие волокна. Эти хроматиновые волокна не концентрируются, а могут находиться либо в компактной форме (гетерохроматин), либо в менее компактной форме (эухроматин). В эухроматине происходят такие процессы, как репликация ДНК, транскрипция и рекомбинация. Во время деления клетки хроматин конденсируется в хромосомы.
Хромосомы — это одноцепочечные структуры, состоящие из конденсированного хроматина. Во время деления клеток посредством митоза и мейоза хромосомы реплицируются для того, чтобы каждая новая дочерняя клетка получила правильное количество хромосом. Дуплицированная хромосома является двухцепочечной и имеет привычную форму X. Две цепи идентичны и соединяются в центральной области, называемой центромерой.
Хроматида — это один из двух клонов дуплицированных хромосом. В конце деления клетки сестринские хроматиды отделяются от дочерних хроматид в новообразованных дочерних клетках.
Разница между хроматином и хромосомой
Определение
Хроматин: Молекулы ДНК в геноме упакованы гистонами, которые образуют хроматин.
Хромосома: самая высокоуложенная структура ДНК формируется в метафазе деления клетки.
период
Хроматин: Хроматин образуется в средней фазе клеточного цикла.
Хромосома: Хромосомы появляются во время метафазы и присутствуют во время анафазы ядерного деления.
Состав
Хроматин: Хроматин состоит из нуклеосом.
Хромосома: Хромосомы конденсированы в хроматиновые волокна.
Condensity
Хроматин: Хроматин конденсируется в 50-кратную двойную спираль нормальной ДНК.
Хромосома: Хромосомы в 10 000 раз более уплотнены, чем обычная двухцепочечная ДНК.
Внешность
Хроматин: Волокна хроматина — это тонкие, длинные, нескрученные структуры.
Хромосома: Хромосомы — это толстые, компактные, похожие на ленту структуры.
Пары
Хроматин: Хроматин представляет собой одиночные непарные волокна.
Хромосома: Хромосома состоит из одной пары.
функция
Хроматин: Хроматин обеспечивает накопление генетического материала в ядре и регулирует экспрессию генов.
Хромосома: Хромосомы обеспечивают правильное расположение генетического материала на экваторе клетки, чтобы генетический материал был равномерно распределен между двумя клетками.
Метаболическая активность
Хроматин: Хроматин обеспечивает репликацию ДНК, экспрессию генов и рекомбинацию.
Хромосома: Хромосомы не обладают метаболической активностью.
подтверждение
Хроматин: Хроматин состоит из двух отростков: Эухроматин и гетерохроматин.
Хромосома: Хромосома обычно гетерохроматическая. Она состоит из таких форм, как метацентрическая, субметацентрическая, акроцентрическая и телоцентрическая.
Визуализация
Хроматин: Хроматин можно наблюдать под электронным микроскопом как структуру с бусинками и нитями.
Хромосома: Хромосому можно наблюдать под световым микроскопом с ее классической четырехрукавной структурой.
Заключение
Хроматин и хромосома — это два типа конденсированных структур молекул ДНК. Интерфазная ДНК находится в форме нитевидной структуры, называемой хроматином. Основная функция хроматина — упаковка двухцепочечной ДНК в ядре клетки с помощью белков гистонов. Молекулы ДНК упакованы в волокна хроматина в 50 раз больше их нормальной структуры. Хромосома появляется только в метафазе клеточного цикла и достигает своей максимально конденсированной структуры. Основная цель хромосомы — обеспечить обмен дубликатами генетического материала между двумя дочерними клетками. Основное различие между хроматином и хромосомой заключается в их конденсации и функционировании во время клеточного цикла.
Справка. Википедия», Н.П.: Фонд Викимедиа, 27 февраля 2017 г., веб. 6 марта 2017 г. 2. Википедия. Н.П. .: Фонд Викимедиа, 3 марта 2017. веб. 6 марта 2017 года.
Изображение предоставлено: 1. «Нуклеосома 1KX5 2», автор Zephyris из Википедии.
Методы исследования хроматина
- ChIP-seq (Секвенирование иммунопреципитации хроматина), направленное против различных модификации гистонов, может использоваться для идентификации состояний хроматина по всему геному. Различные модификации были связаны с различными состояниями хроматина.
- DNase-seq (Секвенирование гиперчувствительных сайтов к ДНКазе I) использует чувствительность доступных участков генома к ДНКаза I фермент для картирования открытых или доступных областей в геноме.
- FAIRE-seq (Секвенирование выделения регуляторных элементов с помощью формальдегида) использует химические свойства связанной с белком ДНК в методе двухфазного разделения для извлечения областей, лишенных нуклеосом, из генома. 26
- ATAC-seq (Анализ для секвенирования транспозиционного доступного хроматина) использует транспозазу Tn5 для интеграции (синтетических) транспозонов в доступные области генома, что, в свою очередь, подчеркивает локализацию нуклеосом и факторов транскрипции в геноме.
- ДНК-след это метод, направленный на идентификацию ДНК, связанной с белками. Он использует маркировку и фрагментацию в сочетании с гель-электрофорезом для идентификации областей генома, которые были связаны белками. 27
- MNase-seq (Секвенирование микрококковой нуклеазы) использует микрококковая нуклеаза фермент для определения положения нуклеосом по всему геному. 2829
- Захват конформации хромосомы определяет пространственную организацию хроматина в ядре, предполагая геномные местоположения, которые физически взаимодействуют.
- Профилирование MACC (Профили доступности микрококковой нуклеазы) использует серию титрования гидролизатов хроматина с микрококковая нуклеаза для определения доступности хроматина, а также для картирования нуклеосом и негистоновых ДНК-связывающих белков как в открытых, так и в закрытых областях генома. 30
Остается загадкой, как деконденсированные хромосомы остаются по существу несцепленными между фазами. Естественно ожидать, что в присутствии топоизомераз ДНК II типа, которые позволяют двуцепочечным участкам ДНК взаимопроникать, все хромосомы должны достичь состояния топологического равновесия. Топологическое равновесие в очень плотных хромосомах между фазами, образующими хромосомные районы, должно приводить к образованию высокоузловатых хроматиновых волокон. Однако методы захвата конформации хромосом (3C) показали, что коллапс контакта геномных расстояний в интерфазных хромосомах практически идентичен состоянию смятых бусин, которое возникает при конденсации длинных полимеров без образования узлов. Для удаления узлов из густонаселенного хроматина требуется активный процесс, который должен не только обеспечить энергию для вывода системы из топологического равновесия, но и направить опосредованные топоизомеразой переходы так, чтобы узлы эффективно расцеплялись и не образовывались. что делает узлы еще более сложными. Было показано, что процесс экструзии хроматиновых петель идеально подходит для активного расцепления хроматиновых волокон в хромосомах между фазами. 31
Хроматин: альтернативные определения
Термин, введенный Уолтером Флеммингом, имеет несколько значений:
- Простое и лаконичное определение: Хроматин — это макромолекулярный комплекс макромолекулы ДНК и макромолекул белка (и РНК). Белки упаковывают и упорядочивают ДНК и контролируют ее функции в ядре клетки.
- Оперативное определение биохимиков: Хроматин — это комплекс ДНК / белок / РНК, экстрагированный из лизированных межфазных ядер эукариот. Какое из множества веществ, присутствующих в ядре, будет составлять часть извлеченного материала, частично зависит от метода, который использует каждый исследователь. Кроме того, состав и свойства хроматина варьируются от одного типа клеток к другому, во время развития определенного типа клеток и на разных стадиях клеточного цикла.
- В ДНК + гистон = хроматин определение: Двойная спираль ДНК в ядре клетки упакована специальными белками, называемыми гистонами. Образованный комплекс белок / ДНК называется хроматином. Основная структурная единица хроматина — нуклеосома.
Первое определение позволяет определить «хроматин» в других областях жизни, таких как бактерии и археи, используя все ДНК-связывающие белки, которые сгущают молекулу. Эти белки обычно называют нуклеоид-ассоциированными белками (NAPs); примеры включают AsnC/LrpC с HU. Кроме того, некоторые археи производят нуклеосомы из белков, гомологичных эукариотическим гистонам. 32
Нобелевские премии
Следующие ученые были удостоены Нобелевских премий за вклад в исследование хроматина:
| Год | ВОЗ | Награда |
|---|---|---|
| 1910 | Альбрехт Коссель (Гейдельбергский университет) | Нобелевская премия по физиологии и медицине за открытие пяти ядерных оснований: Аденин, цитозин, гуанин, тимин и урацил. |
| 1933 | Томас Хант Морган (Калифорнийский технологический институт) | Нобелевская премия по физиологии и медицине за открытия о роли гена и хромосомы в наследственности, основанные на его исследованиях мутации белого глаза у мухи дрозофилы. 33 |
| 1962 | Фрэнсис Крик, Джеймс Уотсон и Морис Уилкинс (Лаборатория молекулярной биологии MRC, Гарвардский университет и Лондонский университет, соответственно). | Нобелевская премия по физиологии или медицине за открытие структуры двойной спирали ДНК и ее значения для передачи информации в живом веществе. |
| 1982 | Аарон Клуг (Лаборатория молекулярной биологии MRC) | Нобелевская премия по химии «За развитие кристаллографической электронной микроскопии и структурное объяснение биологически важных комплексов нуклеиновая кислота-белок». |
| 1993 | Ричард Дж. Робертс и Филип А. Sharp | Нобелевская премия по физиологии «за независимое открытие расщепления генов», при котором участки ДНК, называемые экзонами, экспрессируют белки и прерываются участками ДНК, называемыми интронами, которые не экспрессируют белки. |
| 2006 | Роджер Корнберг (Стэнфордский университет) | Нобелевская премия по химии за открытие механизма, с помощью которого ДНК транскрибируется в информативную РНК. |
Ошибки в организации хроматина
Уплотнение и организация генетического материала имеет решающее значение для различных биологических функций. С дефектами структуры хроматина связано несколько заболеваний, включая Х-сцепленную альфа-талассемию, синдром Рубинштейна-Тейби, синдром Коффина-Лоури, синдром Ретта и другие.

В клетке существует два типа хроматина, которые можно обнаружить с помощью окрашивания: Эухроматин («истинный» хроматин) и гетерохроматин. В первом случае окрашивание слабое, во втором — сильное.
Такая структурная организация ДНК уникальна для эукариот и имеет решающее значение для поведения хромосом и регуляции экспрессии генов.
Если мы оценим пропорции двух типов хроматина в клетке на границе раздела, то обнаружим, что около 90% хроматина — это эухроматин, а остальные 10% — гетерохроматин. Ниже мы подробно опишем каждый тип:
I. Гетерохроматин
Основное различие между двумя типами хроматина связано со степенью конденсации или «упаковки» молекулы на определенных этапах деления клетки.
Хотя может показаться, что генетический материал распределен на границе раздела случайно, это не так.
На этой стадии, когда в ядре наблюдается дифференциальная сегрегация хромосомного материала, организация очень четко выражена.
ДНК хромосом не переплетается с цепочками ДНК других хромосом и остается в определенных регионах, называемых хромосомными областями. Эта организация, по-видимому, способствует экспрессии генов.
Гетерохроматин имеет высокую концентрацию и поэтому недоступен для транскрипционного механизма, поэтому он не транскрибируется. Он также беден генами.
Типы гетерохроматина
Определенные области гетерохроматина сохраняются во всех клеточных клонах, что означает, что они всегда будут вести себя как гетерохроматин. Этот тип гетерохроматина называется конститутивным. Примерами являются конденсированные участки хромосом, центромеры и теломеры.
И наоборот, существуют участки гетерохроматина, которые могут изменять степень своей конденсации в ответ на изменения в паттернах развития или переменных окружающей среды.
Благодаря новым исследованиям эта точка зрения переформулируется, и теперь есть доказательства того, что конститутивный гетерохроматин также динамичен и может реагировать на стимулы.
Хроматин: альтернативные определения
Термин, введенный Уолтером Флеммингом, имеет несколько значений:
- Простое и краткое определение: Хроматин — это макромолекулярный комплекс макромолекулы ДНК и макромолекул белка (и РНК). Белки упаковывают и упорядочивают ДНК и контролируют ее функции в ядре клетки.
- Рабочее определение биохимиков: Хроматин — это комплекс ДНК / белок / РНК, извлеченный из лизированных эукариотических межфазных ядер. Какое из множества веществ, присутствующих в ядре, будет составлять часть извлеченного материала, частично зависит от метода, который использует каждый исследователь. Кроме того, состав и свойства хроматина варьируются от одного типа клеток к другому, во время развития определенного типа клеток и на разных стадиях клеточного цикла.
- Определение ДНК + гистон = хроматин: ДНК двойная спираль в ядре клетки упакована специальными белками, называемыми гистонами. Образованный комплекс белок / ДНК называется хроматином. Основной структурной единицей хроматина является нуклеосома.
Первое определение позволяет определить «хроматин» в других областях жизни, таких как бактерии и археи, используя все белки, которые связывают ДНК и конденсируют молекулу. Эти белки обычно называют нуклеоид-ассоциированными белками (NAPs); примеры включают AsnC/LrpC с HU. Кроме того, некоторые археи производят нуклеосомы из белков, гомологичных эукариотическим гистонам.
Нобелевские премии
Следующие ученые были удостоены Нобелевских премий за вклад в исследование хроматина:
| Год | Кто | Награда |
|---|---|---|
| 1910 | Альбрехт Коссель (Гейдельбергский университет) | Нобелевская премия по физиологии и медицине за открытие пяти ядерных оснований: Аденин, цитозин, гуанин, тимин и урацил. |
| 1933 | Томас Хант Морган (Калифорнийский технологический институт) | Нобелевская премия по физиологии и медицине за открытие роли гена и хромосомы в наследственности, основанное на исследовании мутации «белый глаз» у мухи-дрозофилы. |
| 1962 | Фрэнсис Крик, Джеймс Уотсон и Морис Уилкинс (Лаборатория молекулярной биологии MRC, Гарвардский университет и Лондонский университет, соответственно). | Нобелевская премия по физиологии или медицине за открытие структуры двойной спирали ДНК и ее значения для передачи информации в живом веществе. |
| 1982 | Аарон Клуг (Лаборатория молекулярной биологии MRC) | Нобелевская премия по химии «за кристаллографическую электронную микроскопию и структурное объяснение биологически важных комплексов нуклеиновая кислота-белок». |
| 1993 | Ричард Дж. Робертс и Филип А. Sharp | Нобелевская премия по физиологии «за независимое открытие расщепленных генов», в которых участки ДНК, называемые экзонами, экспрессируют белки и прерываются участками ДНК, называемыми интронами, которые не экспрессируют белки. |
| 2006 | Роджер Корнберг (Стэнфордский университет) | Нобелевская премия по химии за открытие механизма, с помощью которого ДНК транскрибируется в информативную РНК. |
См. Также
- Последовательность активного хроматина
- Хроматида
- Эпигенетика
- Гистон-модифицирующие ферменты
- Позиционное изменение пестроты
- Солевой хроматин
- Транскрипционный взрыв
Дополнительные источники
- Купер, Джеффри М. 2000. Клетка, 2-е издание, Молекулярный подход. Глава 4.2. Хромосомы и хроматин.
- Corces, V.G. (1995). «Хроматиновые изоляторы. Держим усилители под контролем». Природа. 376 (6540): 462–463. Bibcode : 1995Natur.376..462C. doi : 10.1038 / 376462a0. PMID7637775. S2CID26494996.
- Cremer, T. 1985. Von der Zellenlehre zur Chromosomentheorie: Naturwissenschaftliche Erkenntnis und Theorienwechsel in der frühen Zell- und Vererbungsforschungsforschung deröfrönschungen deröfendehröfen der der der forschungen der forschungen der der forschungen der forschungen der forschungen der forschungen der forschungen der der forschungen der forschungen der forschungen der der forschungen der forschungen der der forschungen der forschungen der forschungen der der forschungen der forschungen der der forschungen der der forschungen der forschungen der forschungen. Springer-Vlg., Berlin, Heidelberg.
- Elgin, S.C.R. (ред.). 1995. Структура хроматина и экспрессия генов, vol. 9. IRL Press, Оксфорд, Нью-Йорк, Токио.
- Герасимова, Т. И.; Корсес, В. Г. (1996). «Граничные и изолирующие элементы в хромосомах». Curr. Мнение. Genet. Dev. 6 (2): 185–192. doi : 10.1016 / s0959-437x (96) 80049-9. ПМИД8722175.
- Герасимова Т. И.; Корсес, В. Г. (1998). «Белки группы Polycomb и Trithorax опосредуют функцию изолятора хроматина». Cell. 92 (4): 511–521. doi : 10.1016 / s0092-8674 (00) 80944-7. PMID9491892. S2CID8192263.
- Герасимова Т. И.; Корсес, В. Г. (2001). «ХРОМАТИНОВЫЕ ИЗОЛЯТОРЫ И ГРАНИЦЫ: ВЛИЯНИЕ НА транскрипцию и ядерную организацию». Анну Рев Жене. 35 : 193–208. doi : 10.1146 / annurev.genet.35.102401.090349. PMID11700282. S2CID22738830.
- Герасимова Т. И.; Byrd, K.; Корсес, В. Г. (2000). «Хроматиновый изолятор определяет ядерную локализацию ДНК В процессе цитирования». Mol Cell. 6 (5): 1025–35. doi : 10.1016 / s1097-2765 (00) 00101-5. PMID11106742.
- Ha, S.C.; Lowenhaupt, K.; Rich, A.; Kim, Y.G.; Ким, К. К. (2005). «Кристаллическая структура соединения между B-ДНК и Z-ДНК показывает два экструдированных основания». Природа. 437 (7062): 1183–6. Bibcode : 2005Natur.437.1183H. doi : 10.1038 / nature04088. PMID16237447. S2CID2539819.
- Поллард, Т., и У. Эрншоу. 2002. Клеточная биология. Saunders.
- Saumweber, H. 1987. Расположение хромосом в межфазных ядрах клетки, стр. 223-234. В W. Hennig (ed.), Structure and Function of Eucaryotic Chromosomes, vol. 14. Springer-Verlag, Берлин, Гейдельберг.
- Sinden, R.R. (2005). «Молекулярная биология: повороты ДНК». Природа. 437 (7062): 1097–8. doi : 10.1038 / 4371097a. PMID16237426. S2CID4409092.
- Ван Холд К.Э. 1989. Хроматин. Нью-Йорк: Спрингер- Верлаг. ISBN0-387-96694-3.
- Ван Холде, К., Дж. Златанова, Г. Аренц и Э. Мудрианакис. 1995. Элементы структуры хроматина: гистоны, нуклеосомы и волокна, с. 1-26. В С.С.Р. Элгина (ред.), Структура хроматина и экспрессия генов. IRL Press в Oxford University Press, Оксфорд.







