Эти два последних варианта содержат все данные, которые мы можем найти в кладограмме, плюс дополнительную информацию. Поэтому они не являются взаимоисключающими, а дополняют друг друга.
Становление современной филогенетики
Одной из основных идей современной физической науки является глобальный эволюционизм. Возможно, к этому лучше подходит афоризм известного теоретика физики XX века И. Пригожина: «Мир не есть бытие, а становление». Идея эволюции формирует мировоззрение большинства современных естествоиспытателей и заставляет их включать исторический фактор в причины разнообразия существующего мира.
В биологии идея эволюции имеет большее значение, чем в любой другой отрасли естественных наук. Это объясняется тем, что материал о разнообразии животных и растений дает наибольшую пищу для размышлений. Не зря возникновение современного эволюционного мировоззрения началось с дарвиновской теории эволюции как объяснения происхождения биологических видов.
Тот факт, что биологическое разнообразие является результатом длительного исторического эволюционного процесса, означает, что невозможно полностью понять причины строения и функционирования живых существ без знания их долгой истории. Такая ситуация делает историческую реконструкцию одной из приоритетных задач современной биологии.
Поэтому неудивительно, что в эволюционной биологии возникла особая ветвь — филогенетика, область деятельности которой — реконструкция форм и закономерностей исторического развития живых существ.
Дарвин
Филогенетика возникла в 1960-х годах, сразу после публикации «Происхождения видов» Дарвина в 1859 году». Само понятие филогении появилось в фундаментальной работе немецкого эволюционного биолога Э. Геккеля «Общая морфология», после чего исторические реконструкции играли центральную роль в биологии до 1920-х годов, а любое исследование животных и растений считалось неполноценным, если оно не сопровождалось изображением их филогенетических предков.
В середине двадцатого века ситуация изменилась. Появившаяся в те годы теория эволюции, известная как синтетическая теория эволюции (СТЭ), была полностью сосредоточена на популяционных процессах. Филогенетика, которая занималась и продолжает заниматься в основном макроэволюцией, была отодвинута на «задний план» эволюционных исследований.
В последней трети двадцатого века интерес к филогенетике вновь значительно возрос. Причины этого будут обсуждаться в соответствующем разделе; здесь же достаточно сказать, что в последние десятилетия эволюционная биология переживает то же явление, что и в конце XIX века, которое получило название «филогенетический взрыв».
В данной статье излагается текущее понимание целей и принципов филогенетики и дается обзор классической филогенетики с момента ее зарождения. Кратко представлены приложения современных филогенетических реконструкций к некоторым другим отраслям биологии — биогеографии, систематике и, в некоторой степени, экологии. В заключении дается наиболее сжатый обзор современного понимания генеалогических отношений между большими группами организмов.
Филогенез и филогенетика
Как уже упоминалось, термин филогения (филогенез) был введен Э. Геккелем в середине 19 века. С помощью этой концепции, ставшей общепринятой, он охарактеризовал как процесс исторической эволюции организмов, так и структуру родственных (филогенетических) связей между ними. Понятие эволюции в его современном историческом значении (до этого оно использовалось для определения индивидуального развития организмов), введенное английским философом Р. Спенсером примерно в то же время, также приобрело популярность.
E. Геккель
Это привело к тому, что понятия филогении и эволюции стали рассматриваться как очень близкие или даже синонимичные. Эта классическая интерпретация, приравнивающая филогенез к эволюции, используется и сегодня и встречается в некоторых современных учебниках. В этой очень широкой трактовке филогенез определяется как пути, закономерности и причины исторического развития организмов. Поэтому при таком широком толковании филогенетика рассматривается как причинно-следственная связь.
В конце девятнадцатого века возникло иное понимание взаимосвязи между филогенией и эволюцией: Первое — это сам процесс исторической эволюции, второе — причины этого процесса. Это позволило нам более строго интерпретировать филогению как процесс возникновения и вымирания групп организмов и их специфических характеристик. Следовательно, изучение механизмов филогенеза, то есть причин появления и/или исчезновения групп организмов и их характеристик, в большинстве случаев не входит в задачи современной филогенетики: эта дисциплина носит в основном описательный характер.
Следует отметить еще одно важное различие между классической и современной интерпретацией филогении.
И.И. Шмальгаузен.
Классическая интерпретация ориентирована на организм: Филогенез понимается как историческая эволюция организмов. Эта идея была четко сформулирована выдающимся отечественным ученым-эволюционистом И.И. Шмальгаузеном, который определил филогенез как цепь последовательных онтогений. Этот взгляд основан на понимании того, что самым важным «достижением» биологической эволюции является организм как наиболее полная биологическая система.
В настоящее время ведется активная работа над биоцентрическим пониманием природы филогенеза. Она основана на идее, что биологическая эволюция — это самоэволюция среды обитания как интегрированной системы, и одним из аспектов этой эволюции является филогенез.
Такое понимание биологической эволюции в целом и филогенеза в частности более соответствует современному пониманию общих законов эволюции, разработанному наукой синергетикой. Ее основы были заложены И. Пригожиным, основателем теории динамики неравновесных систем (за что он был удостоен Нобелевской премии). Одной из особенностей этой динамики является структурирование этих систем в процессе их развития: появление все большего числа элементов, которые объединяются в комплексы разной степени общности. Биосфера является типичной неравновесной системой; следовательно, ее развитие, обычно называемое биологической эволюцией, можно рассматривать как процесс ее структурирования (биосферы).
В этом отношении одним из важнейших результатов эволюции является глобальная структура живого мира Земли, которая проявляется в многоуровневой иерархии групп, интегрированных и организованных по-разному. Эту структуру можно приблизительно представить как двухкомпонентную, состоящую из двух основных иерархий, каждая из которых является результатом определенных физических, биологических и, в некоторой степени, исторических процессов.
Онтогенез и филогенез
В видеоуроке на примере лютеинового и постэмбрионального развития объясняются типы онтогенеза, периоды онтогенеза и стадии эмбрионального развития. В этом уроке рассматриваются следующие термины: Онтогенез, зигота, зигота, бластогенез, нуклеация, бластула, гастрогенез, нейрула, нейрогенез, филогенез.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этим и другим видеоурокам комплекта, вам необходимо добавить его в свой личный кабинет.

2. Распространите видеоуроки в своих личных кабинетах среди учеников.
3. Смотрите статистику просмотра видеоуроков учениками.
Конспект урока «Онтогенез и филогенез»
Процесс индивидуального развития человека от начала его существования до конца жизни называется онтогенезом.
У бактерий и простейших онтогенез практически идентичен клеточному циклу.
У многоклеточных видов, размножающихся бесполым путем, онтогенез начинается, когда группа клеток из родительского организма выделяется и затем делится путем митоза, образуя новую особь со всеми ее системами и органами.
У видов, размножающихся половым путем, онтогенез начинается в момент оплодотворения яйцеклетки и образования зиготы — первой клетки нового организма.
У животных можно выделить три типа онтогенеза: личиночное, овариальное и внутриутробное развитие.
Тип личинкиРазвитие.
Он встречается, например, у насекомых, рыб и земноводных. Желток в яйцах этих организмов маленький, и зигота быстро развивается в личинку, которая питается и развивается самостоятельно. Через некоторое время происходит метаморфоз, т.е. превращение личинки во взрослую особь.
Некоторые виды даже проходят цепную трансформацию от личинки до взрослой особи.
У кузнечика, например, личинка питается, развивается и мутирует несколько раз. После окончательного появления на свет он становится взрослой особью с крыльями и дальше не развивается.
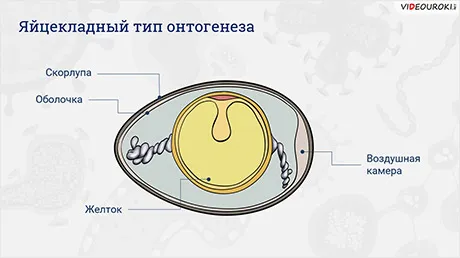
Тип яйцекладаОнтогенез
Он наблюдается у рептилий, птиц и яйцекладущих млекопитающих, яйца которых богаты желтком. Эмбрион этих видов развивается внутри яйца; личиночная стадия отсутствует.
Эмбрион питается желтком. Содержимое яйца окружено двумя оболочками — внутренней и внешней. Внешняя часть — это оболочка. После откладки яйца на тупом конце яйца постепенно образуется воздушная камера.
Однако у большинства млекопитающих яйцеклетка не формируется, и эмбрион развивается в организме самки в репродуктивном органе — матке.
Тип эндометрияОнтогенез
Это происходит у большинства млекопитающих, включая человека. Развивающийся эмбрион остается в организме матери и образует временный орган — плаценту, через которую организм матери удовлетворяет все потребности развивающегося эмбриона: Дыхание, питание, выделение и т.д. Внутриутробное развитие завершается процессом рождения.
Периоды онтогенеза
Любой тип индивидуального развития у многоклеточных животных принято делить на два периода: эмбриональный и постэмбриональный.
Эмбриональный периодначинается с момента оплодотворения и представляет собой процесс формирования сложного многоклеточного организма, в котором присутствуют все системы органов.
Этот период заканчивается вылуплением личинки из оболочек (при личиночном типе онтогенеза), вылуплением особи из яйца (при овариальном типе) или рождением особи (при внутриутробном типе).
Постэмбриональная фазаначинается с завершением эмбрионального периода. Она включает в себя половое созревание, подростковый возраст, взрослую жизнь и старение, которое заканчивается смертью.
Стадии эмбрионального развития на примере копья.
Следует помнить, что ланцетовидныйеЛанцетник — примитивное морское животное семейства ланцетников. Ланцетник считается промежуточным звеном между позвоночными и беспозвоночными.
Что такое филогенетическое дерево?
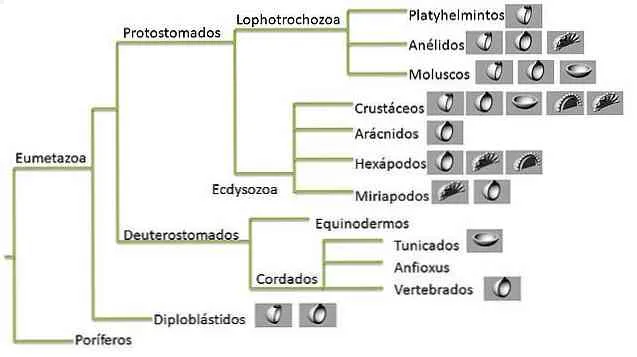
Филогенетическое дерево — это графическое представление эволюционной истории группы организмов. Эта схема исторических взаимоотношений и есть филогения, которую пытаются оценить исследователи.
Деревья состоят из узлов, соединенных «ветвями». Конечные узлы каждой ветви представляют собой конечные таксономические группы и представляют последовательности или организмы, для которых известны данные — это могут быть живые или вымершие виды.
Внутренние узлы представляют собой предполагаемых предков, а предок в корне дерева представляет собой предка всех последовательностей, представленных в графе.
Как интерпретируются филогенетические деревья?
Существует множество способов представления филогенетического дерева. Следовательно, важно знать, обусловлены ли различия, наблюдаемые между двумя деревьями, разной топологией, т.е. реальными различиями, соответствующими двум графам, или просто способом их представления…..
Например, порядок появления меток в верхней части может меняться без изменения смысла графического представления, обычно это название вида, рода, семейства и других категорий.
Это объясняется тем, что деревья похожи на мобильный телефон, в котором ветви могут вращаться, не меняя взаимосвязи между представленными видами.
В этом смысле не имеет значения, как часто меняется порядок или как часто вращаются «плавающие» объекты, потому что способ их соединения не меняется — и это все, что имеет значение.
Как реконструируются филогении?
Филогенез — это гипотеза, основанная на косвенных доказательствах. Размывание филогенеза похоже на работу следователя, идущего по следу на месте преступления.
Биологи часто формулируют свои филогении, используя знания из различных дисциплин, таких как палеонтология, сравнительная анатомия, сравнительная эмбриология и молекулярная биология…..
Ископаемая летопись, хотя и неполная, дает очень ценную информацию о времени происхождения видов.
Со временем молекулярная биология обогнала все вышеперечисленные области, и большинство филогенетических исследований проводятся на основе молекулярных данных…
Цель реконструкции филогенетического дерева имеет ряд важных недостатков. Существует около 1,8 миллиона видов, которые были названы, и еще много видов, которые пока не описаны.
И хотя значительное число ученых ежедневно пытаются реконструировать взаимоотношения между видами, у нас все еще нет полного дерева.
Гомологичные персонажи
Когда биологи хотят описать сходство между двумя структурами или процессами, они могут сделать это в терминах общего происхождения (гомология), аналогии (функция) или гомоплазии (морфологическое сходство).
Для реконструкции филогении используются только гомологичные признаки. Гомология является ключевым понятием для эволюции и реконструкции отношений между видами, поскольку только она адекватно отражает общее происхождение организмов….
Предположим, мы хотим вывести филогению трех групп: Птицы, летучие мыши и люди. Для достижения нашей цели мы решили использовать верхние конечности в качестве признака, который помогает нам различать структуру отношений.
Поскольку птицы и летучие мыши имеют структуры, которые могут летать, мы можем ошибочно заключить, что летучие мыши и птицы являются более близкими родственниками, чем летучие мыши и люди. Почему мы пришли к неправильному выводу? Потому что мы использовали похожий, а не гомологичный знак.
Чтобы найти правильное родство, мне приходится искать гомологичные признаки, например, наличие волос, молочных желез и трех маленьких косточек в среднем ухе. Гомологии, однако, нелегко диагностировать.
Основные принципы построения филогений
Вероятно, самым важным правилом в филогенезе сегодня является принцип дихотомии: предполагается, что из трех таксонов два должны быть более родственными, чем третий. Поэтому филогении обычно напоминают дихотомически разветвленные деревья. Когда порядок ветвления определен для всего дерева, оно считается полностью решенным. Иногда в филогенетических исследованиях встречаются «кусты» или политомии, то есть места, где порядок ветвления неясен, и в этом случае говорят, что дерево неполностью разрешено. Этот принцип несовершенен, поскольку развитие таксонов не всегда дихотомично. Если оказывается, что эта дихотомия не соответствует реальным фактам, исследователи обращаются к другим системам, таким как филогенетические сети 8.
Еще в 1960-х годах методы создания филогений делились на два основных направления — фенетическое и кладистическое. В то время анализ родства основывался на морфологических признаках 12. Даже когда в построение филогении включаются молекулярные признаки, основные принципы анализа родства остаются по сути теми же.
- В фенетике построение филогении основано на общем сходстве двух видов — то есть, чем больше общих признаков, тем ближе они друг к другу;
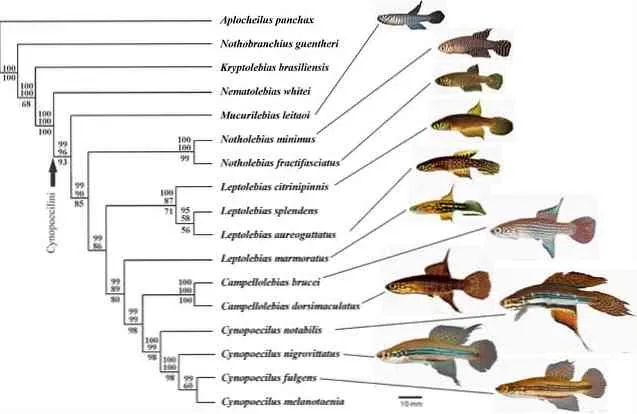
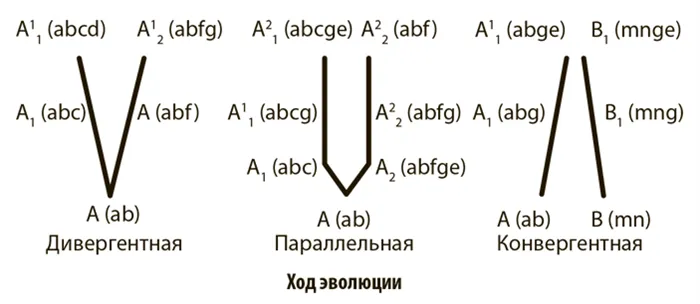
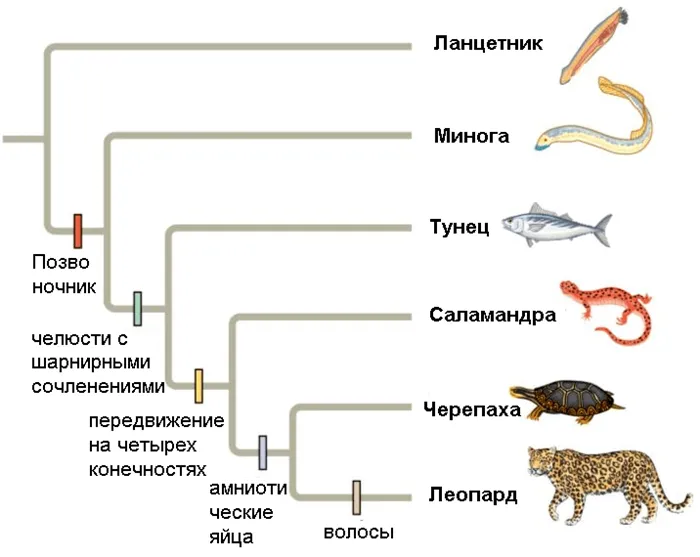
- В кладистике же считается, что только уникальные для какой-либо группы признаки можно использовать для оценки родства таксонов. Родоначальником кладистического анализа является немецкий ученый Вилли Хенниг 6. Этот автор также ввел и терминологию, которая широко используется до сих пор. Уникальные признаки называются апоморфиями ; ветви, которые объединяются апоморфиями — это клады ; а сама филогения называется кладограммой (рис. 2) 12.
Чтобы проиллюстрировать это, представьте себе три вида животных: домовую мышь, сумчатого и кенгуру. Домовая мышь и сумчатая мышь очень похожи внешне, но сумчатая мышь и кенгуру имеют общий порок развития — оболочку, что позволяет предположить родство этих двух видов. Но, конечно, филогенетический анализ основан на гораздо большем количестве признаков, и группы могут иметь более одного порока развития.
Первые шаги. ДНК—ДНК гибридизация
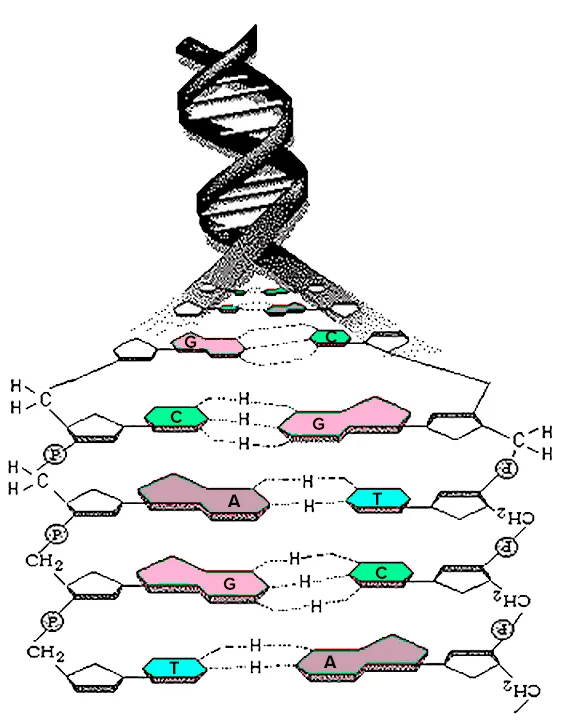
Первые попытки использовать ДНК в качестве основы для построения семейного древа жизни были феноменальными. В 1984 году американские ученые Сибли и Алкист 13 предприняли первую попытку использовать ДНК для выяснения филогении различных видов приматов. Они использовали метод, называемый гибридизацией ДНК-ДНК. Метод основан на том, что мутации постоянно возникают во время репликации ДНК. Это приводит к тому, что даже два близких родственника имеют разные последовательности ДНК, не говоря уже о видах. Другими словами, чем дальше организмы находятся на семейном древе, тем сильнее различаются их ДНК. В этом методе молекулы ДНК двух видов смешиваются друг с другом таким образом, чтобы образовать «гибридные» двойные спирали, где одна половина принадлежит одному виду, а другая — другому. Затем эти «гибриды» нагревают, и исследователь изучает, при какой температуре двойная спираль расщепляется (или разделяется) на две части. Считается, что чем выше температура, необходимая для расщепления «гибрида», тем сильнее связь между молекулами ДНК двух разных видов, и, следовательно, тем ближе становятся виды (рис. 3).
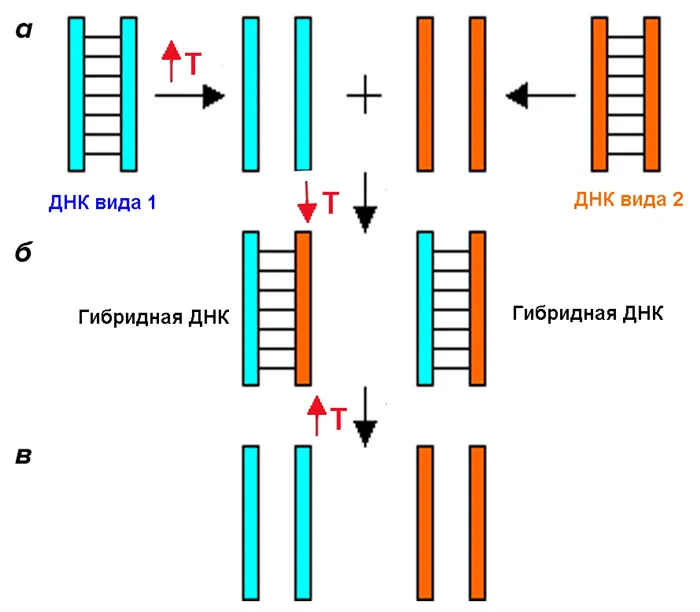
Вскоре стало ясно, что этот метод не может быть очень точным. Дело в том, что гены могут гибридизировать не только со своими гомологичными генами (ортологичные гены), но и с копиями этих генов, которых в геноме может быть очень много (парологичные гены). С развитием методов секвенирования генов последовательности ДНК или белков, хранящиеся в компьютерных файлах, стали основным источником для построения филогений. В последние годы скорость накопления генетической информации продолжает расти, и филогения окончательно утвердилась в качестве метода анализа и обработки биологических текстов.
Метод матрицы расстояний (distance matrix)
Метод матрицы расстояний является по сути феноменологическим методом. Он основан на вычислении парных различий между соответствующими генами всех видов, участвующих в данном анализе. Это делается следующим образом: Гены каждого исследуемого вида сравниваются в каждой позиции нуклеотида, и чем больше различий найдено, тем больше «расстояние» между видами. Затем создается таблица, в которую вводится это значение для каждой возможной пары сравниваемых генов. Таблица расстояний является исходным материалом для алгоритмов построения деревьев.
Наиболее популярным из этих алгоритмов является метод ближайшего соседа. Среди проанализированных видов были обнаружены два вида с наименьшими различиями в последовательности (т.е. наиболее схожие). С помощью таблицы данные этих видов «объединяются», а затем анализируются в едином виде. Виды проходят через этот процесс один за другим, пока не будет найдено единое, полностью разрешенное дерево. Этот алгоритм хорош тем, что он относительно прост и подходит для обработки больших массивов данных (рис. 4) 3.
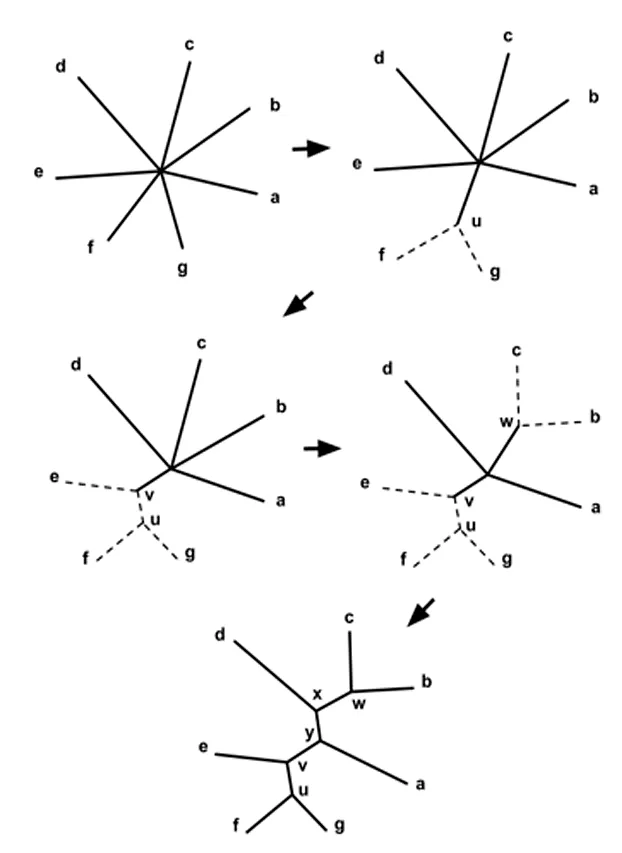
Однако различные авторы указывают на некоторые недостатки метода ближайшего соседа. Например, считается, что этот метод хуже работает для филогенетически удаленных таксонов 4, 17. Другим недостатком является то, что метод всегда дает дерево только с одним возможным порядком ветвления 3. Причина в том, что алгоритм строит филогению без сравнения с другими, в то время как кладистические методы оценивают деревья с разными порядками ветвления. Хотя сегодня методы матрицы расстояний редко используются в серьезном филогенетическом анализе, они применяются, например, для быстрого построения филогений близкородственных бактерий и вирусов 18.
Параллельная эволюция
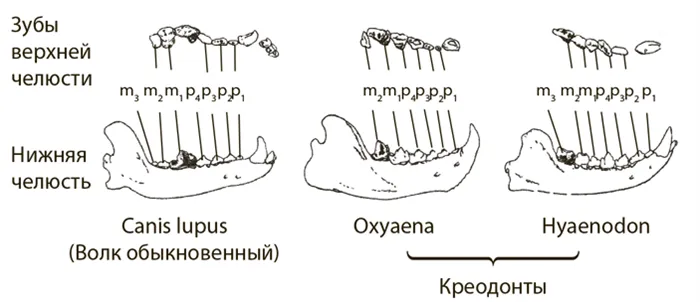
После разделения адаптивной зоны на подзоны, когда дивергенция прекращается, каждый таксон продолжает адаптироваться к условиям своей новой подзоны. Затем следует период параллельной эволюции. Подобные адаптации формируются на общей основе, унаследованной от предков. Это объясняется тем, что подзоны являются частью общей адаптационной зоны и условия жизни в них в основном одинаковы. Замечательный пример параллелизма можно найти у неправильных морских ежей.
Морские ежи, как и морские звезды, являются хищниками. Их скелет состоит из известковых пластин с пятиугольной симметрией, которые вместе образуют изогнутую вверх полусферу. Рот находится на нижней плоской стороне полушария, а анус — в середине изогнутой верхней стороны. В юрском периоде нерегулярные морские ежи были приспособлены к питанию илом. Главное преимущество грязи в том, что ее много. Здесь много органического материала для питания, но также много грязи. Питание илом требовало заглатывания большого количества этого субстрата, что удлиняло пищеварительный тракт. У пяти групп морских ежей это вызвало смещение ануса назад. В результате тело (скелет) изменило форму и стало более вытянутым. Образовались неправильные морские ежи (Рисунок 54).
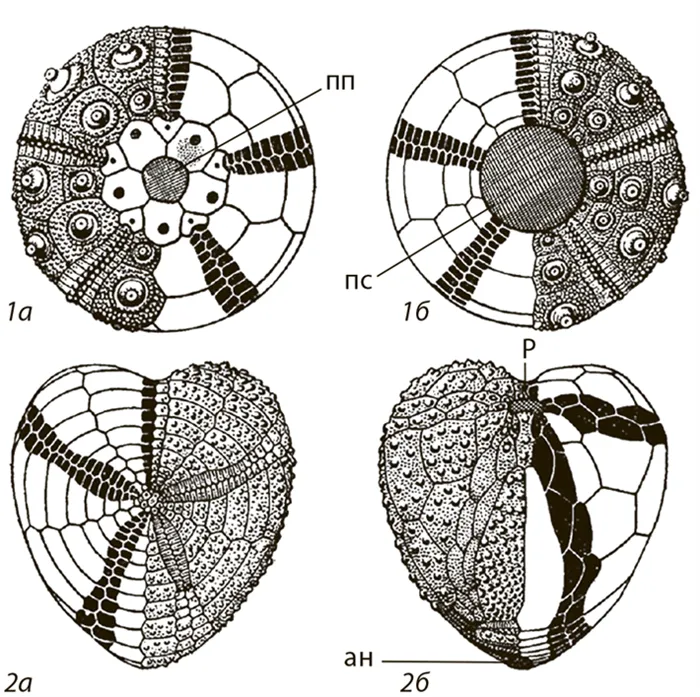
Рисунок 54. схематическое изображение строения панциря правильного ( 1 ) и неправильного ( 2 ) морских ежей (по Druschitz, 1974) (a — вид сверху, b — вид снизу): an — анальное отверстие; np — перипростернальный (небольшое поле, в центре которого находится анус); ps — брюшина с ротовым отверстием; p — рот.
Чем более схожи адаптивные зоны или подзоны, тем более схожи параллельно развивающиеся признаки. Например, панцирные позвонки — вы можете видеть их в рыбе на вашем столе — произошли от двуногих рыб, от костных ганноидов (малоизвестной группы саланоидов), от костных рыб и от амфибий. Самая глубокая параллель обнаруживается в эволюции организации тканей. Позвоночные и насекомые имеют полосатую структуру мышц. В более примитивном состоянии, без развитых мышечных волокон, он существует даже у медуз. Великий отечественный историк А.А. Заварзин показал глубочайшую параллель в эволюции нервной ткани, когда сравнил первичные и вторичные организмы: полихет и личинок миндалин, насекомых и позвоночных.
Эти параллели усложняют филогенетические реконструкции и часто приводят к ошибкам в них. Неполнота палеонтологической летописи приводит к тому, что организмы из разных параллельных таксонов включаются в один и тот же филогенетический ряд. Сходство между ними большое, но отношения отдаленные.
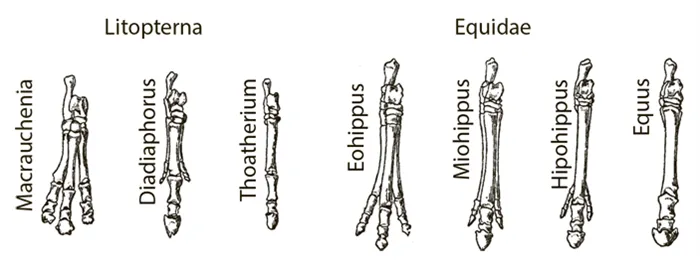
Обратите внимание, что параллели не всегда развиваются одновременно. В нескольких случаях было показано, что в одной группе они развиваются раньше, чем в другой. Затем они пишут о «филогенетических предсказаниях», «пророческих группах» и т.д. Эти факты охотно используются противниками эволюции путем естественного отбора. Если два таксона приобретают одинаковые признаки, один раньше, а другой позже, отбор здесь ни при чем. Другие законы эволюции, как утверждается, направляют эволюцию по заранее определенному пути.
Примером асинхронного (не одновременного) параллелизма является эволюция непарнокопытных Litopterna в Южной Америке и лошадей, включая зебр и ослов. Литоптерны, как и лошади, были солипедами. Эволюция обеих групп происходила одинаково. Боковые пальцы ног укорачивались до тех пор, пока не оставался только средний палец и копыто. Lithoptera существовали с конца палеоцена до их вымирания в плейстоцене. Эволюция современных лошадей началась в эоцене.
Конвергенция
Конвергенция — это появление сходных признаков у неродственных таксонов. В одинаковых условиях окружающей среды естественный отбор приводит к эволюции сходных адаптаций. При конвергенции адаптация к абиотическим факторам более очевидна. Чтобы скользить от дерева к дереву, выгодно увеличить площадь воздушной опоры — кожные складки по бокам тела и конечностей расширяются. У водных позвоночных форма тела оптимальна (рис. 57).
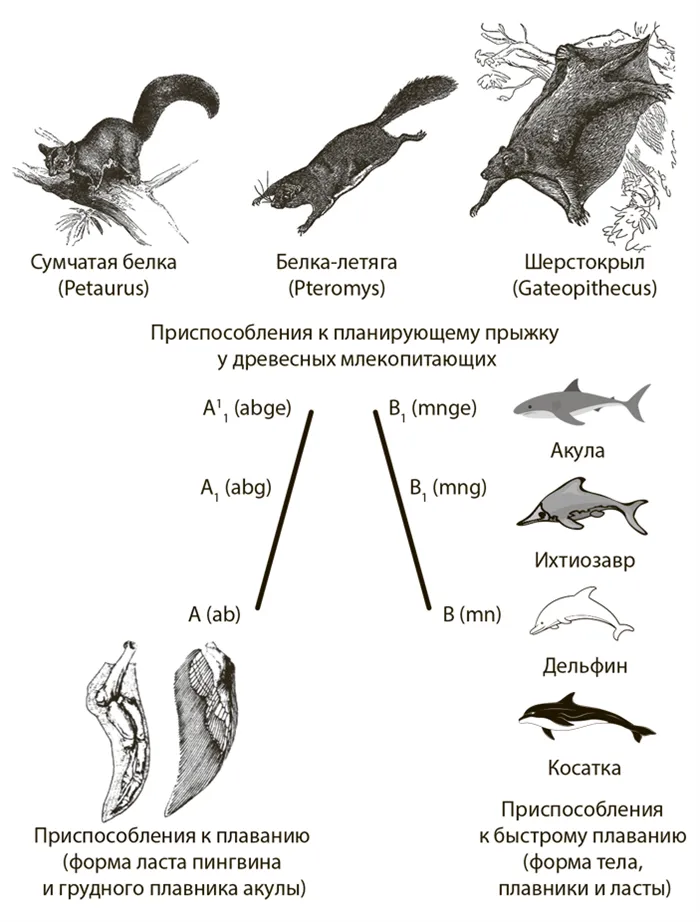
Рисунок 57. примеры конвергентной эволюции у позвоночных (по Schmalhausen, 1969).
Когда они тесно связаны, трудно отличить конвергенцию от параллелизма. У крупных хищников развились очень длинные клыки в верхней челюсти, длиной до 20 см. Известно, что более десяти видов «сабельных зубов» существовали в разное время, от 50 миллионов до 10 000 лет назад. Такие же клыки Saburotooth использовал Barburofelis, микенское животное, которое не принадлежало к семейству кошачьих, хотя и было его дальним родственником. В Южной Америке до объединения с Северной Америкой обитали зауродонтовые тилакосмилы — сумчатые, у которых также были зауродонтовые клыки. И Barburofelis, и Tylacosmilus имели костяные лопасти в нижней челюсти, своего рода ножны для длинных клыков верхней челюсти.
Хотя существуют различные мнения о том, как спатодонты используют свои зубы, ясно, что это охотничья адаптация, возникшая в результате биотических взаимодействий, а не абиотических факторов. В семействе кошачьих это асинхронные и аллопатрические варианты. Саблезубый тигр — это конвергенция. Предположение о том, что клыки Barburofelis являются результатом конвергенции или параллелизма, неясно.
Сабертуты были крупными хищниками. Их скелеты хорошо изучены. В более старых и менее хорошо изученных формах легко спутать конвергенцию с параллелизмом, а параллелизм — со стадией в филогении рассматриваемого таксона. Как вы уже знаете, реконструкция эволюции таксонов — это работа поколений исследователей, исправляющих ошибки своих предшественников.
Монофилия и полифилия в происхождении надвидовых таксонов
Существуют две противоположные точки зрения на происхождение таксономических групп более высокого порядка — подотрядов, порядков, надпорядков. Ученые, отстаивающие теорию монофилии, предполагают, что каждый таксономический вид более высокого ранга является потомком одного исходного вида. Все разнообразие низших систематических групп является результатом адаптивной радиации.
Сторонники многовидовой теории предполагают, что таксоны высокого ранга могут происходить от нескольких родительских групп, которые даже систематически удалены друг от друга.
Существует также промежуточный вариант — концепция широкой монофилии: происхождение считается монофилетическим, если порядок дочернего таксона выше или равен порядку таксона происхождения, независимо от того, сколько филогенетических линий привело к дочернему таксону. Концепция ограниченной монофилии, отстаиваемая некоторыми палеонтологами, в которой дочерний таксон возникает из нескольких одновременно эволюционирующих таксонов более низкого порядка, очень близка к этой концепции.
Примером широкой монофилии является схема происхождения наземных позвоночных, предложенная выдающимся шведским палеонтологом Э. Ярвиком (рис. 58). Порядок Ripidistia, порядок Cystaceae, который является прототипом для высшего порядка четвероногих — тетрапод (наземных позвоночных), состоял из двух подотрядов. Согласно этой схеме, хвостатые являются потомками одного из подотрядов, а все остальные — потомками другого. Бесхвостые амфибии, рептилии и даже предки млекопитающих самостоятельно переползали из воды на сушу. Только птицы уже отделились от рептилий на суше. Ярвик считал себя сторонником полифонии. Согласно теории широкой монофилии, надпорядок тетрапод превосходит порядок Ripidistia и весь порядок Cystoptera.
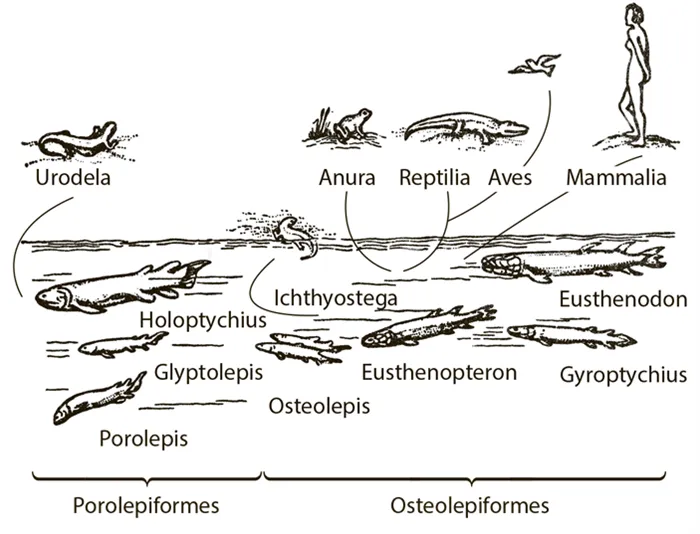
Рисунок 58. Происхождение наземных позвоночных (из Jarvik 1970).
Дебаты о поли- или монофилии имеют два аспекта: таксономический и эволюционный.
Систематическая теория требует физической системы, основанной на строгой монофилии. Если выясняется, что части таксономической группы, связанные внешними признаками, на самом деле имеют разное происхождение, таксономическая группа разделяется. Так было и с грызунами, которых пришлось разделить на грызунов и кроликов. Птицы с длинной шеей и длинным клювом делились на журавлей, аистов и т.д.
Эволюционная теория, с другой стороны, не запрещает независимое внедрение различных таксонов в одну и ту же зону адаптации. Примеры включают водных млекопитающих, от китов до моржей, тюленей до морских котиков, морских выдр, каланов.
Вопрос не в том, могут ли таксоны разного происхождения обитать в одной и той же зоне адаптации или нет. Могут — хорошим примером является расселение морских млекопитающих в Мировом океане. Это вопрос определения происхождения таксона или таксонов, которые колонизируют территорию. Этот вопрос не возникает у водных млекопитающих. Различие между китами, моржами, тюленями и серыми тюленями не вызывает споров. В более сложных случаях требуется детальное сравнение сходств и различий. В последние десятилетия все чаще используются молекулярно-биологические данные. Они были использованы, например, для сокращения подотрядов нелетающих птиц, таких как страусы, кассауары, бантамы и актинииды, до подотрядов.







