Зрительный нерв окружен несколькими мембранами. Межкохлеарное пространство заполнено спинномозговой жидкостью. Эта жидкость, спинномозговая жидкость, постоянно находится в движении и направляется в третий желудочек (полая структура, относящаяся к среднему мозгу и расположенная в пространстве между зрительными долями).
Размножение клеток: Мейоз
Библиографическая ссылка: Сазонов В.Ф. Размножение клеток: мейоз. электронный ресурс // Кинезиолог, 2009-2021: сайт. Дата обновления: 28.04.2021. URL: http://kineziolog.su/content/razmnozhenie-kletok-meyoz (дата обращения: __.__.20___). ___________ Мейоз, стадии. Гаметогенез.
Определение понятия
Мейоз — это специфическое деление двух клеток, которое происходит путем сокращения хромосомного набора с двойного диплоидного до одинарного гаплоидного, и при котором хромосомы правильно распределяются между четырьмя дочерними клетками. © 2015-2021 Сазонов В.Ф. © 2015-2021 kineziolog.su .
Обычным результатом мейоза у животных являются гаметы, т.е. гаметы с одним гаплоидным набором хромосом вместо двойного диплоидного набора, характерного для соматических клеток. У растений, однако, результатом мейоза являются не гаметы, а споры. Однако в этом случае споры также имеют гаплоидный набор хромосом. Это означает, что процесс редукции приводит к гаплоидии.
По этой причине процесс мейоза используется, например, у мхов. В результате мейоза из диплоидных форм мхов образуются гаплоидные споры. Конечно, споры не являются гаметами, но понятие мейоза остается типичным и для мхов: это двойное деление диплоидной клетки, приводящее к образованию четырех гаплоидных клеток (только в данном случае это спора, а не гамета). Споры не могут быть оплодотворены, они должны сначала вырасти во взрослое растение (гаплоид), которое производит гаметы (путем митоза, а не мейоза).
Так, у мхов гаметы гаплоидных взрослых растений производятся из соматических клеток, которые уже гаплоидны, поэтому сперматозоиды и яйцеклетки производятся путем митоза, но не мейоза.
Так, у растений мейоз дает начало спорам, а митоз — гаметам (половым клеткам). Это означает, что растения используют мейоз и митоз «наоборот» по сравнению с животными.
Метафорически можно сказать, что мейоз — это «решающий» процесс! Она начинается сразу с решения стоящей перед ней задачи, то есть уже с предлога первого разделения двух. И эта задача — формирование набора хромосом из двойного. Далее мы опишем, что именно происходит в префазе 1.
Чтобы получить одинарный, а не двойной набор хромосом, во время муссирования происходит два последовательных деления. Более того, основная задача получения одинарного набора хромосом вместо двойного решается уже во время первого деления. Это не «половинки» хромосом, как при митозе, а парные гомологичные хромосомы, которые распределяются по двум дочерним клеткам. По этой причине первое деление мейоза называется редукцией, так как при нем общее число хромосом уменьшается вдвое. Второе редукционное деление (изомерное деление) решает другую проблему: оно делит каждую хромосому на две сестринские хроматиды и распределяет эти хроматиды поровну между двумя клетками.
Как лучше запомнить и понять формулы, обозначающие общее число хромосом на разных стадиях мейоза?
Расшифровка формул хромосомного набора
1n — единичный набор хромосом (1 — единичный, n — общий).
testis.jpg
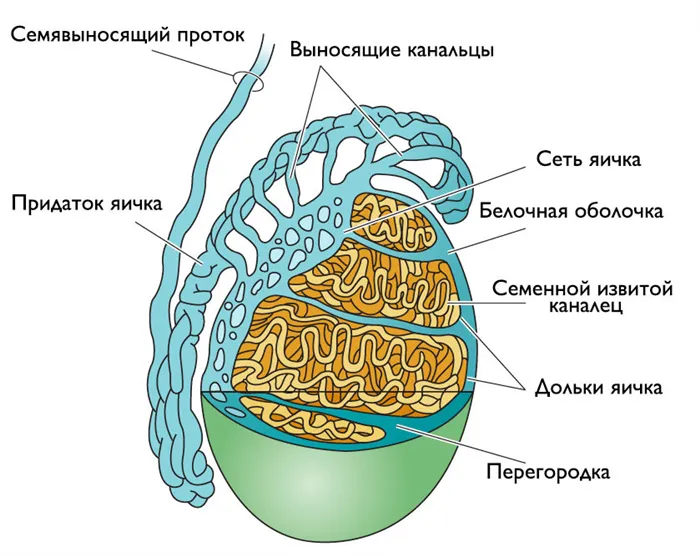
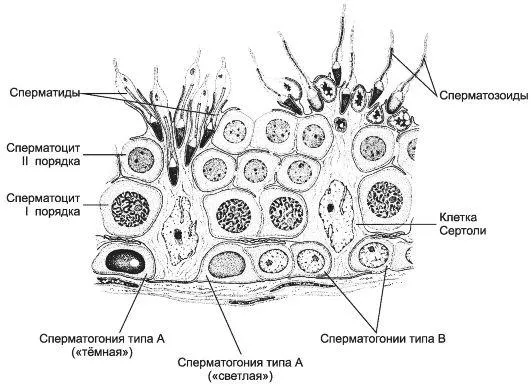
Рисунок 1. Сперматогенез
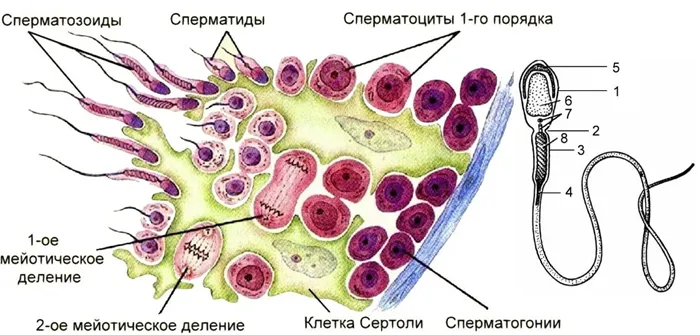
Рисунок 2. Сперматогенез.
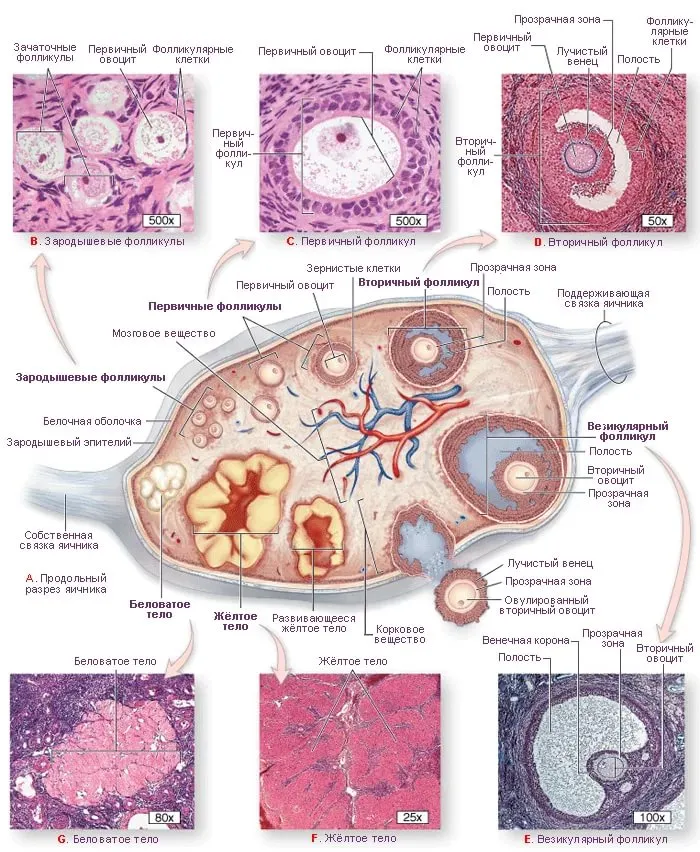
Рисунок: Оогенез (овуляция).
Структура и функция синаптонемного комплекса
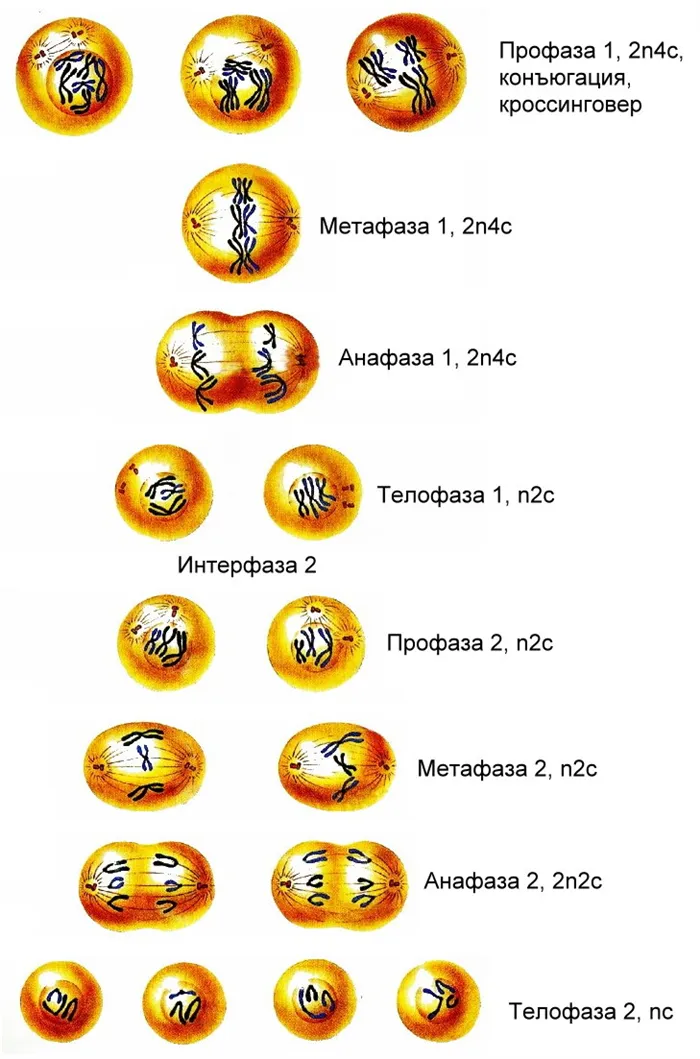
Рисунок 2. Очень тонкий продольный срез синаптоневрального комплекса свиньи Ascaris suum, состоящего из двух латеральных и одного центрального элемента. Плотно конденсированный хроматин расположен вблизи латеральных элементов. Центральный элемент состоит из перекрывающихся белковых «зубцов молнии». Фото автора. Изображение: Природа
Синаптонный комплекс состоит из двух белковых осей гомологичных хромосом, соединенных белковой «молнией» (Рисунок 2). Зубцы молнии представляют собой бактероидные димеры параллельных и эквилатерально ориентированных белковых молекул с длинной α-спиралью в середине молекулы. У дрожжей S. cerevisiae это белок Zip1, у млекопитающих и человека — SCP1 (SYCP1). Эти белки прикреплены своими С-концами к осям хромосом (боковые элементы комплекса), а их N-концы направлены друг к другу в центральном пространстве (рис. 3). Заряженные «шпоры» — чередующиеся пики плотности положительных и отрицательных зарядов аминокислот (рис. 4), комплементарное взаимодействие которых обеспечивает прочную электростатическую связь зубцов, расположены на N-концах молекул.
Так называемое центральное пространство комплекса (промежуток между осями белков, который заполнен зубцами фиксатора и имеет ширину около 100 нм) и весь комплекс (его поперечное сечение составляет около 150-200 нм) невозможно увидеть под обычным световым микроскопом, поскольку весь комплекс покрыт хроматином. Впервые синаптонный комплекс наблюдался в чрезвычайно тонких (толщиной 0,8 мкм) срезах семенников рака и мыши с помощью просвечивающего электронного микроскопа. Он был открыт в 1956 году двумя американскими исследователями, М. Мозесом и Д. У. Фоссета 2, независимо друг от друга.
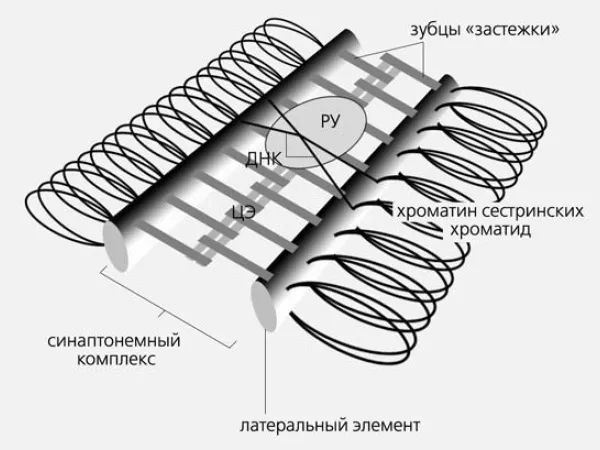
Рисунок 3. Схематическое изображение структуры синаптонового комплекса у дрожжей, дрозофилы и млекопитающих. CE — центральный элемент; RU — узел рекомбинации, в котором были экспериментально идентифицированы ферменты, участвующие в сшивании. Рисунок: Природа
В настоящее время для изучения комплекса используется так называемый метод микродиссекции. Клетки яичка (или растительного эфира) после гипотонического шока помещают на пластиковую подложку на предметном стекле микроскопа. Содержимое препаровальной камеры фиксируют слабым раствором формальдегида и смешивают с солями тяжелых металлов (предпочтительно AgNO3Стекло исследуется под фазово-контрастным микроскопом, и клетки, которые должны содержать комплекс, отбираются путем непрямого обнаружения. Кружок пленки, содержащий нужную клетку, собирается на металлической сетке и помещается на электронный микроскоп (рис. 5). При необходимости клетки обрабатывают антителами против интересующих белков перед окрашиванием. Эти антитела помечены калиброванными шариками коллоидного золота, которые хорошо видны под электронным микроскопом.
Белки синаптонемного комплекса
В 1970-х годах мы получили косвенные доказательства того, что синаптосомный комплекс формируется путем самоорганизации его элементов, которая может происходить и без хромосом. Сама природа организовала этот эксперимент, а мы смогли его наблюдать. Оказалось, что у свиных аскарид пакеты или «стопки» совершенно правильно расположенных морфологических элементов синаптического комплекса появляются в цитоплазме клеток, готовящихся к мейозу I (хотя хромосом в цитоплазме нет: они находятся в ядре). Поскольку в ядрах клеток на стадии подготовки клеток к мейозу отсутствует синаптонный комплекс, была выдвинута гипотеза о неполном контроле последовательности мейотических событий в этом примитивном организме. Избыток вновь синтезированных белков в цитоплазме приводит к их полимеризации и возникновению структуры, не похожей на синаптонный комплекс 3. Эта гипотеза была подтверждена только в 2005 году работой международной группы исследователей из Германии и Швеции. Они показали, что когда ген, кодирующий белок молнии млекопитающих (SCP1), вводится в соматические клетки, выращенные в искусственной питательной среде, и они активируются, внутри культивируемых клеток возникает мощная сеть белков SCP1, «застегивающихся» подобно центральному пространству комплекса. Образование слоя непрерывных белковых молний в культуре клеток означает, что предсказанная способность сложных белков к самосборке была продемонстрирована 4.
В 1989 и 2001 годах О.Л. Коломиец и Ю.С. Федотова из нашей лаборатории изучали физическую «разборку» синаптонных комплексов на последних стадиях их существования. Этот многоступенчатый процесс лучше всего демонстрируется в материнских клетках пыльцы пыльников ржи, где происходит частичная синхронизация мейоза 5. Было установлено, что боковые элементы комплекса деградируют путем постепенного «разматывания» белковой суперспирали, которая имеет три уровня упаковки (рис. 6).
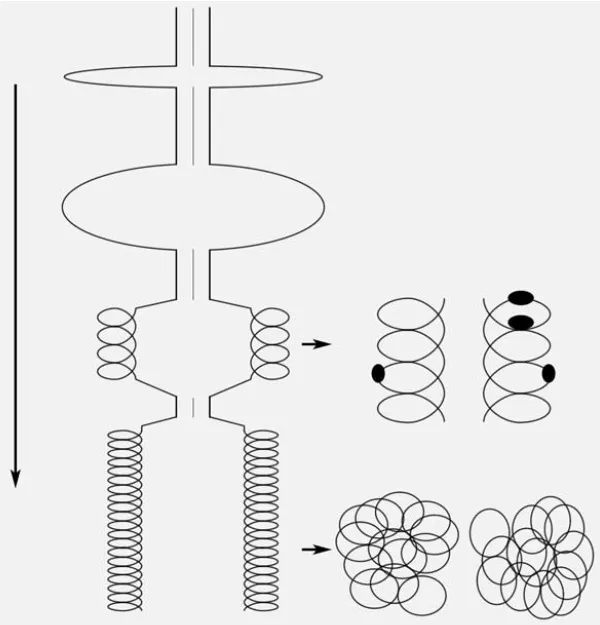
Рисунок 6. Этапы деградации синаптонового комплекса во время фазы дупликации мейоза у ржи. Все линии показывают только белковую нить, которая постепенно удлиняется. Рисунок: Природа
Основой удлиненных латеральных элементов является комплекс из четырех белков когезина (cohesin). Накануне миоза на хромосомах появляется специфический белок когезин Rec8, который заменяет соматический когезин Rad21. Затем он соединяется с тремя другими когезиновыми белками, которые также присутствуют в соматических клетках, но вместо соматического когезина SMC1 появляется муцин-специфический белок SMC1b (его N-концевой конец отличается на 50% от N-концевого конца соматического белка SMC1). Этот когезиновый комплекс расположен внутри хромосомы между двумя сестринскими хроматидами и удерживает их вместе. Специфические для мейоза белки ассоциируются с комплексом когезина, который становится основным белком хромосомных аксонов и превращает их (эти аксоны) в латеральные элементы синаптосомного комплекса 6. У млекопитающих основными белками синаптосомного комплекса являются SCP2 и SCP3, у дрожжей — белки Hop1 и Red1, а Rec8 — муцин-специфический белок.
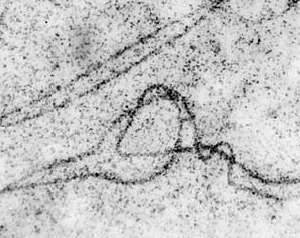
Рисунок 7. Фрагмент синаптонемного комплекса, демонстрирующий гетерозиготную хромосомную перестройку — инверсию, которая выглядит как типичная петля инверсии. Фото. Изображение: Природа
В целом, это хорошая фундаментальная наука, но можете ли вы использовать ее в практических целях? Вы можете. В середине 1980-х годов британские исследователи и наша лаборатория показали на нескольких экспериментальных моделях, что микроскопия синаптонных комплексов позволяет обнаружить в два раза больше хромосомных перестроек (делеций, сдвигов, инверсий), чем традиционный метод анализа хромосом на стадии метафазы (рис.). 7): Синаптонный кластер — это скелетная структура мейотических хромосом в профазе. На этом этапе хромосомы примерно в десять раз длиннее, что значительно повышает разрешение анализа. Однако рассмотреть клубок хромосом в профазе практически невозможно, а жесткие скелетные структуры синаптосомного комплекса не боятся разматываться, более того, электронный микроскоп может обнаружить мини-хромосомы, недоступные для светового микроскопа.
Нам стало интересно, можно ли определить причину бесплодия у потомства облученных мышей, исследуя комплекс синаптома, а не хромосомы. Было обнаружено, что у бесплодных мышей, унаследовавших хромосомные сдвиги от своих родителей, эти перестройки обнаруживались комплексом в 100% исследованных клеток, тогда как обычные методы «метафазного» анализа выявляли их только в 50% клеток 13. Испанская исследовательская группа обследовала более 1000 мужчин, страдающих бесплодием. У трети из них причину бесплодия не удалось определить заранее, а при исследовании комплекса синаптонома из клеток яичек этих пациентов диагноз удалось поставить половине из них: Причиной бесплодия является отсутствие синаптономного комплекса, что приводит к тому, что сперматоциты (клетки-предшественники сперматозоидов) не развиваются, т.е. процесс мейоза и весь сперматогенез «заторможен» 14. Аналогичные результаты были получены O. L. Коломиец вместе с врачами из Харькова. Исследование синаптонового комплекса в сочетании с другими аналитическими методами повысило частоту выявления причин бесплодия у обследованных пациентов мужского пола с 17 до 30% 15. Некоторые английские клиники активно использовали такие методы уже в 1990-х годах. Конечно, такая диагностика требует высоких теоретических и практических навыков врачей и использования электронных микроскопов. За исключением Института общей генетики имени С.И. Вавилова Российской академии наук, российские лаборатории еще не достигли этого уровня. Н.И. Вавилова РАН (Москва) и Институт общей генетики СО РАН (Новосибирск).
Можно подумать, что интенсивное изучение механизмов мейоза неизбежно приведет к применению полученных знаний в областях биологии и медицины, связанных с фертильностью живых существ, включая человека. Однако закон научного применения неопровержим: бессмысленно что-либо «заставлять». Задача профессионалов — следить за научными разработками и использовать их. Именно такого подхода придерживаются ведущие фармацевтические и биотехнологические компании.
С момента открытия мейоза (1885) до открытия синаптонового комплекса (1956) прошло около 70 лет, а с 1956 до открытия белков синаптонового комплекса (1986) — еще 30 лет. В последующие 20 лет мы узнали о структуре этих белков, генах, кодирующих их, взаимодействии белков при сборке и функционировании синаптонных комплексов, особенно об их взаимодействии с белками ферментов рекомбинации ДНК и т.д., то есть больше, чем за предыдущие 30 лет описательных цитологических исследований. Для расшифровки основных молекулярных механизмов мейоза, вероятно, потребуется не более двух десятилетий. История науки, как и вся цивилизация, характеризуется «сжатием времени», все большим сжатием событий и открытий.
Симптомы глиомы хиазмы
Глиома хиазмы развивается в основном или как следствие распространения глиомы зрительного нерва на зрительный бугорок. На практике доказано, что глиома хиазмы распространяется вдоль зрительного нерва и инвазирует орбиту, с риском развития в гипоталамусе или полости третьего желудочка.
Очень часто глиома хиазмы может сопровождаться реактивным арахноидитом, приводящим к образованию агглютинированных структур или субарахноидальных кист.
По своим клиническим проявлениям глиома хиазмы отличается от всех других глазных опухолей направлением роста и локализацией.
Наиболее важные симптомы хиазмальной глиомы:
- снижение остроты зрения;
- видоизменение зрительных полей;
- ликворно-гипертензионный синдром;
- эндокринно-обменные сбои.
Ухудшение зрения вызвано тем, что опухоль давит на зрительный нерв. Другой причиной может быть разрушение зрительных нервов, в которое вовлечена развивающаяся глиома.
По мере прогрессивного роста хиазматической глиомы в направлении зрительного нерва обычно наблюдается утолщение инфильтрированных волокон зрительного нерва. Это приводит к пережатию нерва в области канала зрительного нерва.
Нарушение зрения в этом случае можно объяснить двумя способами. Возможно, что острота зрения снижается на ранней стадии развития глиомы хиазмы. Нарушение зрения является односторонним, а нарушение зрения на втором глазу проявляется только через некоторое время (от нескольких месяцев до нескольких лет).
Главной особенностью хиазматической глиомы является ее медленное, но прогрессирующее развитие, которое проявляется в асимметричном снижении зрения пациента. Когда глиома распространяется на орбиту, у пациента развивается прогрессирующая форма экзофтальма в дополнение к снижению остроты зрения.
Какие изменения происходят в поле зрения, зависит в основном от расположения самой опухоли. Чаще всего глиома располагается в передней части зрительного пути, так что поражаются оптические волокна вплоть до их перехода на другую, противоположную сторону. В этом случае наблюдается двулучепреломление, связанное с сужением поля зрения как одного глаза, так и другого. Не исключено концентрическое сужение поля зрения с частичной потерей зрения. Если глиома хиазмы расположена в заднем отделе (за слиянием зрительных нервов), возникает гомонимная гемианопсия, то есть выпадение отдельных половин поля зрения.
Другие симптоматические явления, связанные с развитием глиомы хиазмы и способствующие ее инвазии в область гипоталамуса, включают в себя
- гиперсомнию;
- ожирение;
- гипоталамический синдром;
- диэнцефальный синдром;
- диабет несахарного типа;
- гиперкортицизм;
- нарушение в психике;
- раннее половое созревание.
Другими симптомами, сопровождающими хиазманглиому, являются тошнота, характерная острая головная боль и ощущение тяжести вокруг глазных яблок. Все эти симптомы обусловлены реактивным арахноидитом. Симптомы становятся более тяжелыми, когда глиома инвазирует полость третьего желудочка, что приводит к обструктивным гидродинамическим нарушениям с дальнейшим прогрессированием до гидроцефалии.
Диагностика глиомы хиазмы
Поскольку первым и наиболее важным клиническим проявлением глиомы хиазмы является нарушение зрения, пациент обычно сначала обращается к офтальмологу. Специалист назначит офтальмологическое обследование, которое включает тесты для определения остроты зрения, а также компьютерную периметрию и простое обследование глазного яблока.
- Периметрия — один из самых важных методов исследования, который помогает выяснить степень поражения зрительных путей, получив четкое представление о месторасположении глиомы.
- Как известно, на начальном этапе своего формирования, глиома хиазмы симптоматически никак себя не проявляет. При дальнейшей офтальмоскопии могут быть выявлены застойные диски (зрительных нервов), а также первые признаки атрофии зрительного нерва первичного типа. Посредством исследования зрительных ВП врач устанавливает уровень пораженного участка зрительного нерва.
- Полученные результаты комплексного исследования у офтальмолога, указывающие на присутствие обширного процесса зрительных путей внутри черепа, будут поводом для получения консультации у невролога с дальнейшим исследованием головного мозга через МРТ или КТ.
- Если не представляется возможным проведения КТ или МРТ головного мозга, тогда врачом назначается прохождение рентгенографии черепа с целью выявления деформации турецкого седла, которое при глиоме хиазмы приобретает грушевидную форму.
- В дополнение может быть проведен рентген орбит (косая проекция). Если же глиома хиазмы прорастает в плоскость орбиты, то рентген указывает на соответствующее расширение зрительных отверстий, которое может быть одно- или двусторонним, и составлять около 9 мм.
Томографические исследования глиомы хиазмы
Однако имейте в виду, что КТ или МРТ намного точнее рентгеновского снимка. С помощью методов томографии можно определить наличие глиомы, ее расположение, степень инвазии в зрительные нервы и тип роста. Также можно определить, могла ли опухоль проникнуть в ткани головного мозга (или гипоталамуса).
Основным недостатком томографии является то, что она не может отличить глиомы хиазмы от других глиом аналогичного расположения, так как на их месте может появиться тератома или ганглионеврома. Дифференциальный диагноз может быть поставлен только при гистологическом исследовании полученных опухолевых клеток, которые обычно получают при стереотаксической биопсии.







