Организм (обычно диплоидный = 2n) называется полиплоидным, если он имеет один или более полных наборов хромосом. В отличие от точечных мутаций, инверсий и дупликаций хромосом, это крупномасштабный процесс, т.е. он происходит в полных наборах хромосом.
Что такое полиплоидия, ее значение и роль в образовании видов
Полиплоидия — это множественное число хромосомных наборов в клетках растений или животных, кратное числу хромосом в одной клетке.
Гаметы обычно гаплоидны (имеют только один набор хромосом), тогда как соматические клетки диплоидны. Если клетки живого существа содержат более двух наборов хромосом, его называют полиплоидным. Триплоиды имеют 3 набора, тетраплоиды — 4, а пентаплоиды — 5. Особи с нечетным числом хромосом не могут дать потомство. Это происходит потому, что их гаметы не имеют полного набора хромосом и не могут делиться.

Как возникает полиплоидия
Полиплоидия — это одна из форм изменчивости. Она обеспечивает биоразнообразие, когда потомство приобретает новые признаки, фенотипически отличающиеся от признаков родителей.
Наиболее важным условием является то, что во время редукции не происходит расхождения хромосом. В этом случае гамета имеет диплоидный набор хромосом. Скрещивание с гаплоидной клеткой приводит к образованию триплоидной клетки, а слияние клеток с одинаковым количеством хромосомных наборов приводит к образованию тетраплоидной зиготы.
У каких организмов наблюдается полиплоидия? У диких видов растений, особенно цветковых, полиплоидия широко распространена (около половины видов являются полиплоидными). Поскольку растения могут размножаться вегетативно, полиплоидия не мешает им производить потомство, в отличие от животных.
У животных это явление встречается редко, поскольку нерасхождение хромосом во время мейоза приводит к генетическим ошибкам. Полиплоидия у животных характерна для некоторых гермафродитов (червей) и особей, размножающихся без оплодотворения. Для большинства простейших характерно большое количество хромосомных наборов (около сотни).
Роль полиплоидии в образовании видов
Около 75 % современных сортов растений являются полиплоидными. К ним относятся овощи и фрукты, зерновые, цитрусовые и лекарственные растения. Популярными триплоидами являются арбузы и виноград без косточек. Эти виды демонстрируют стерильность триплоидных организмов, поскольку не могут производить потомство.
Полиплоидия нашла применение в садоводстве при создании новых сортов растений. Метод основан на искусственном увеличении числа хромосом в клетках живых организмов, которое всегда кратно гаплоидному набору. В результате происходит интенсивный рост клеток и человека в целом.
На сегодняшний день выведено много новых, высокоурожайных и устойчивых сортов. Для достижения желаемого эффекта используется мутаген, такой как колхицин. Он предотвращает расхождение хромосом во время деления.
Мутации с увеличением числа хромосом также происходят под воздействием температуры, радиации или вследствие изменения внутреннего состояния клетки. Таким образом, под воздействием внешних факторов веретено деления не формируется, и процесс распределения генетической информации между дочерними клетками останавливается. Интрамиелинизация может быть причиной полиплоидии — количество хромосом удваивается, но само ядро клетки не делится.
Клеточная полиплоидия делает растения более устойчивыми к изменениям окружающей среды и воздействию чужеродных веществ. Эта устойчивость обусловлена тем, что при гибели многих гомологичных хромосом большинство из них продолжают функционировать.
Аллополиплоидные организмы также используются для размножения. Хромосомные наборы этих особей различаются: набором генов, формой или количеством хромосом. Например, при скрещивании растений разных родов, таких как рожь и пшеница, получается гибрид с одним набором ржи и одним набором пшеницы. Потомство больше не сможет воспроизводить себе подобных; только увеличение числа хромосом у обоих растений позволит возобновить репродуктивную функцию.
Автополиплоидия
Автополиплоидия — это размножение гаплоидных хромосомных наборов одного вида. Первая мутация, автотетраплоид, была описана в начале двадцатого века. G. de Friese в Oenothera. У него было 14 вместо 7 пар хромосом. Дальнейшее исследование числа хромосом у представителей различных семейств выявило широкое распространение автополиплоидии в растительном мире. При автополиплоидии происходит либо равномерное (тетраплоид, гексаплоид), либо избыточное (триплоид, пентаплоид) увеличение числа хромосом. Автополиплоиды отличаются от диплоидов большим размером всех органов, включая репродуктивные. Это связано с увеличением размеров клеток за счет повышения плюрипотентности (ядерно-плазменного индекса).
Растения по-разному реагируют на увеличение числа хромосом. Когда в результате полиплоидии число хромосом превышает оптимальное, аутополиплоиды, хотя и демонстрируют некоторые признаки гигантизма, в целом более слаборазвиты, как, например, пшеница с 84 хромосомами. Автополиплоиды часто проявляют некоторую степень стерильности из-за аномалий мейоза во время созревания половых клеток. Иногда высокополиплоидные формы оказываются нежизнеспособными и полностью бесплодными.
Аутополиплоидия является результатом нарушения процесса деления клеток (митоза или мейоза). Митотическая полиплоидия возникает, когда дочерние хромосомы не делятся во время профазы. Если это происходит во время первого деления зиготы, все клетки эмбриона являются полиплоидными; на более поздних стадиях образуются соматические мозаики — организмы, части тела которых состоят из полиплоидных клеток. Митотическая полиплоидизация соматических клеток может происходить на разных стадиях онтогенеза. Мейотическая полиплоидия возникает, когда во время образования половых клеток мейоз пропускается или заменяется митозом или другим типом нередукционного деления. Это приводит к образованию нередуцирующих гамет, слияние которых приводит к образованию полиплоидного потомства. Такие гаметы чаще всего образуются у апомиктических видов и только в исключительных случаях у половых.
Очень часто автотетраплоиды не скрещиваются с диплоидом, от которого они произошли. Если скрещивание между ними проходит успешно, образуются автотриплексы. Нередкие полиплоиды обычно чрезвычайно стерильны и не могут быть размножены семенами. Однако у некоторых растений триплоидия, по-видимому, является оптимальным уровнем богатства. Эти растения имеют признаки гигантизма по сравнению с диплоидами, например, триплоидные тополя, триплоидная сахарная свекла и некоторые сорта яблок. Триплоидные формы размножаются либо подражанием, либо вегетативным размножением.
Для искусственного получения полиплоидных клеток используется сильный яд Colchicum automnale, действие которого поистине универсально: полиплоиды можно получить из любого растения.
Аллополиплоидия
Аллополиплоидия — это дублирование набора хромосом у отдаленных гибридов. Например, если гибрид имеет два разных генома AB, то полиплоидный геном будет ABA. Межвидовые гибриды часто бывают стерильными, даже если скрещиваемые виды имеют одинаковое число хромосом. Это происходит потому, что хромосомы разных видов не гомологичны, и поэтому процессы спаривания и расхождения хромосом нарушены. Нарушение еще более выражено, когда номера хромосом не совпадают. Если в яйцеклетке гибрида происходит спонтанная дупликация хромосом, он становится аллополиплоидным и содержит два диплоидных набора родительского вида. В этом случае мейоз протекает нормально, и растение становится плодовитым. Такие аллополиплоиды С.Г. Навашин назвал амфидиплоидами.
В настоящее время известно, что многие полиплоидные формы, встречающиеся в природе, являются результатом аллополиплоидии, например, обыкновенная пшеница с 42 хромосомами является амфидиплоидом, возникшим в результате скрещивания тетраплоидной пшеницы и диплоидного родственного вида Aegilops (Aegilops L.) и последующего удвоения всех хромосом триплоидных гибридов.
Аллополиплоидия была обнаружена у многих видов сельскохозяйственных культур, таких как табак, рапс, лук, ива и др. Таким образом, аллополиплоидия у растений является одним из механизмов видообразования наряду с гибридизацией.
Анеуплоидия
Анеуплоидия — это изменение числа отдельных хромосом в кариотипе. Возникновение анеуплоидии является результатом неправильного расхождения хромосом в процессе деления клеток. Анеуплоиды часто возникают в потомстве автополиплоидов, в которых гаметы с аномальным числом хромосом образуются из-за неправильного расхождения поливалентов. Если одна гамета имеет хромосомный набор из n + 1, а другая — из n, в результате слияния образуется трисомия — диплоид с одной лишней хромосомой в наборе. Когда гамета с хромосомным набором n — 1 сливается с нормальной гаметой ( n ), в результате получается моносома — диплоид с отсутствующей одной хромосомой. Если в наборе отсутствуют две гомологичные хромосомы, организм называется нуллисомным. У растений часто жизнеспособны как моносомы, так и трисомы, хотя потеря или добавление хромосомы вызывает некоторые изменения в фенотипе. Последствия анеуплоидии зависят от количества хромосом и генетического состава дополнительной или утраченной хромосомы. Чем больше хромосом в наборе, тем меньше растения подвержены анеуплоидии. Трихомы растений несколько менее жизнеспособны, чем нормальные особи, и их фертильность снижается.
Моносомы у сельскохозяйственных культур, таких как пшеница, часто используются в генетическом анализе для определения расположения различных генов. В пшенице, а также в табаке и других растениях были созданы моносомные линии, состоящие из линий, в каждой из которых была потеряна одна хромосома из нормального набора. У пшеницы также известны нулевые изомеры с 40 хромосомами (вместо 42). В зависимости от того, какая из пар хромосом 21a отсутствует, их жизнеспособность и фертильность ограничены.
Анеуплоидия у растений тесно связана с полиплоидией. Это хорошо видно на примере синих трав. В пределах рода Роа известны виды, образующие полиплоидные ряды с числом хромосом, кратным главному числу (n = 7): 14, 28, 42, 56. В траве проса прерийного эуплоидия почти исчезла и сменилась анеуплоидией. Число хромосом у разных биотипов этого вида составляет от 50 до 100 и не является кратным основному числу, что связано с анеуплоидией. Анеуплоидные формы сохраняются, поскольку размножаются партеногенетически. У растений анеуплоидия рассматривается генетиками как один из механизмов эволюции генома.
У животных и людей изменение числа хромосом имеет гораздо более серьезные последствия. Примером моносомии является дрозофила, у которой отсутствует 4-я хромосома. Это самая маленькая хромосома в наборе, но она содержит ядерный организатор и, таким образом, образует ядро. Его отсутствие приводит к уменьшению размеров мухи, снижению плодовитости и изменению некоторых морфологических признаков. Однако мухи жизнеспособны. Потеря гомолога из других пар хромосом приводит к летальному исходу.
У человека геномные мутации обычно приводят к тяжелым наследственным заболеваниям. Например, моносомия на Х-хромосоме приводит к синдрому Шерешевского-Тернера, который характеризуется физическим, умственным и половым недоразвитием у носителей этой мутации. Трисомия на Х-хромосоме имеет аналогичный эффект. Наличие в кариотипе дополнительной 21-й хромосомы приводит к развитию хорошо известного синдрома Дауна (более подробно об этом рассказывается в лекции «Генетика человека»).
Читайте другие темы в книге «Генетика и репродукция. Теория. Задания. Ответы» :
Полиплоидия
Полиплоидия — это состояние, при котором общее число хромосом увеличивается в несколько раз по сравнению с гаплоидным. Это состояние часто встречается в природе, например, такие растения, как виноград, сахарная свекла, водоросли, грибы и т.д. являются полиплоидными. Однако у людей полиплоидия является серьезным генетическим заболеванием с высоким уровнем смертности.
Почему развивается полиплоидия?
У человека диплоидный набор хромосом, 2n, является врожденным. Диплоидные клетки образуются в результате слияния гаплоидных клеток во время оплодотворения. Дочерняя клетка получает один набор от материнской клетки и родительской клетки. Этот паттерн может быть нарушен различными патологическими состояниями, некоторые из которых будут упомянуты здесь:
- нарушение расхождения хромосом в процессе митоза;
- слияние соматических клеток либо их ядер;
- оплодотворение одной яйцеклетки двумя сперматозоидами;
- нарушение нормального течения мейоза.
Риск полиплоидии возрастает, когда клетки подвергаются воздействию физических или химических агентов, которые повреждают веретено деления.
Методы диагностики полиплоидии
Цитогенетическое исследование служит для изучения всех хромосом пациента (кариотипа). Она осуществляется в несколько этапов:
- получение биологического материала. Обычно это кровь пациента;
- выделение из образца необходимого количества лимфоцитов;
- культивация клеток и последующая обработка митогеном. Данное вещество запускает процесс митоза;
- фиксация материала, его окрашивание и микроскопирование.
Наконец, врач может увидеть все хромосомы в окуляре микроскопа и рассмотреть их.
Различные цитогенетические исследования позволяют выявить не только полиплоидию, но и ряд других заболеваний (моносомия, трисомия, делеция, инверсия и т.д.), а также оценить размер и форму хромосом.
Вы можете пройти диагностику своих генетических заболеваний в Медицинском геномическом центре Genomed.
Хромосомные перестройки и мейоз
Первыми событиями, которые ученые обнаружили в полиплоидных клетках с помощью микроскопа, могли быть хромосомные перестройки. Большинство этих аномалий (особенно во время мейоза) приводят если не к смерти, то, как правило, к бесплодию. Однако нельзя сказать, что эти изменения не важны для эволюции. Некоторые из них могут привести к радикальным фенотипическим изменениям, которые могут потребоваться в случае резкого изменения окружающей среды («сальтаторное» возникновение нового вида) 16.
В основе различных хромосомных перестроек лежит один и тот же механизм гомологичной рекомбинации 17. Другими словами, все участки ДНК, демонстрирующие гомологию, могут рекомбинировать: как внутри генома, так и между отдельными геномами в полиплоиде. Если в диплоидном ядре рекомбинация обычно не мешает правильному расхождению гомологичных хромосом, то в полиплоидном ядре образуются поливалентные хромосомы, состоящие из гомологичных и гомологичных (происходящих из разных геномов) хромосом. Этот процесс похож на вязание косичек — двойных, тройных и т.д. Чем больше их переплетено, тем меньше вероятность того, что коса распутается. На уровне хромосом это проявляется в неадекватном расхождении, например, когда все четыре хромосомы четверки оказываются в одной гамете и ни одной в другой. В результате возникает хромосомная нестабильность, что крайне нежелательно для организма. И даже если дивергенция происходит, она чревата всевозможными дефектами: удалениями, сдвигами, разворотами и т.д.
Как мы можем избежать этого и обеспечить спаривание и расхождение только гомологичных хромосом, исключив из этого процесса гомологичные хромосомы? Для этого природа создала генетический механизм, подавляющий гомологичную рекомбинацию. У пшеницы, например, этот механизм контролируется локусом Ph1, который содержит ряд генов с аналогичными функциями 18. Продукты этих генов изменяют гистоновые белки хромосом. Наличие этого локуса (Ph1+) приводит к выработке дефектных белков, в результате чего снижается модификация гистонов и задерживается распад хромосом перед первым редукционным делением. Затем происходит синапсис хромосом с последующей рекомбинацией.
Как мы знаем, конденсация хромосом необходима для формирования синапса и образования комплекса синапса. Теперь вернемся к процессу плетения. Для правильного плетения пряди должны быть одинаковой длины. Если они неровные или между ними большое расстояние, ничего не получится. Аналогичным образом, спаривание хромосом в преддверии мейоза зависит от их сходства — чем больше сходство (а в случае связей — чем больше), тем больше вероятность того, что они спарятся первыми. Если процесс конденсации или выравнивания длины хромосом под влиянием локуса Ph1 ограничен во времени, то в спаривании будут участвовать в основном гомологи (рис. 3).
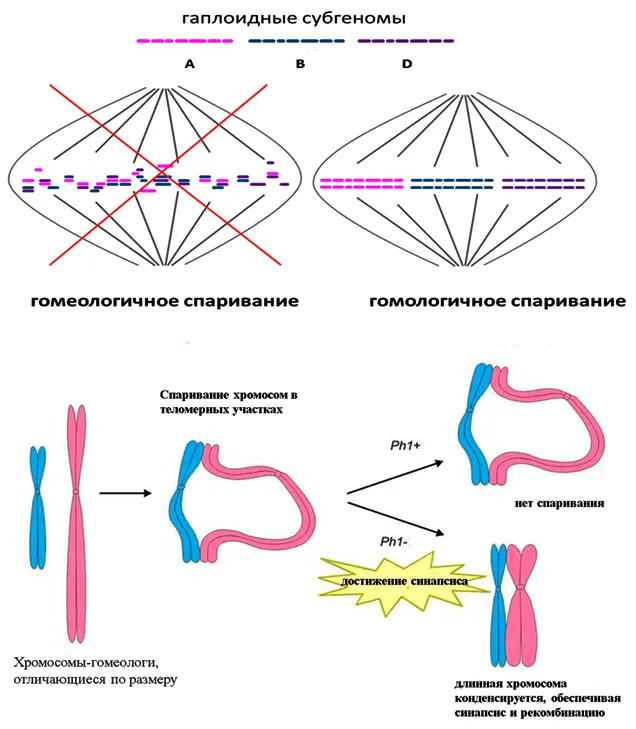
Здесь мы подходим к очень важному условию сосуществования двух или более геномов в одном ядре. Чем больше структурных различий накапливается на хромосомах, тем больше шансов преодолеть цитогенетическую несовместимость во время мейоза. Это звучит парадоксально, поскольку их генетические системы и функциональные механизмы должны оставаться схожими. Иначе все закончится, как в известной саге о лебеде, раке и щуке. Чтобы избежать этого, необходим дифференциальный подход, то есть геном может накапливать различия только в той части генома, которая не столь необходима для его функционирования и воспроизводства.
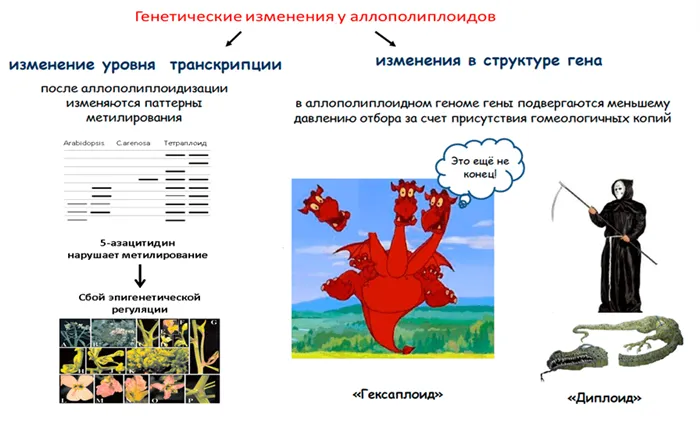
Транскриптомные изменения и феномен «ядрышкового доминирования»
Что касается кодирующей части генома, то все значительные перестройки неприемлемы по очевидным причинам. Однако нельзя сказать, что в этой части ничего не происходит. Наиболее распространенным и самым ранним эффектом аллополиплоидии является изменение транскрипционной активности генов (транскрипционные изменения) 20. Эти изменения могут вызвать транспозоны (см. выше), хотя это не обязательно. Наиболее ярким примером является феномен ядерного доминирования.
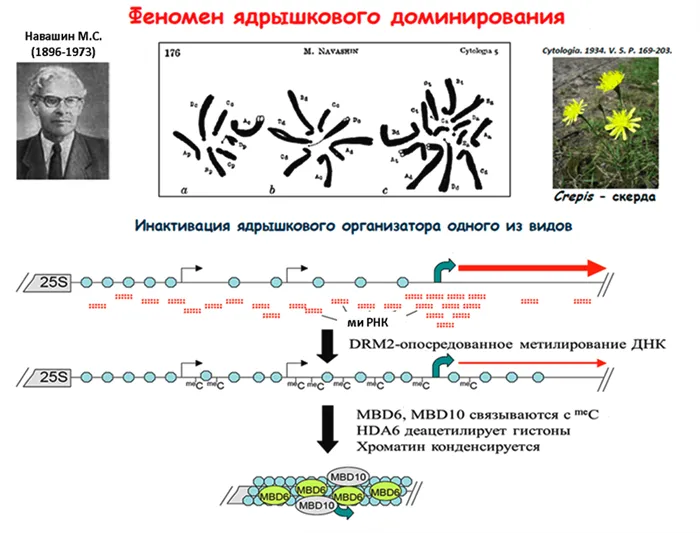
Сначала несколько слов о ядрах. Это цитологические структуры хромосом, в которых собираются субъединицы рибосом; они состоят из генов, кодирующих рибосомальную РНК (рРНК). Оказывается, ядро — это своего рода фабрика рибосом, где количество генов рРНК определяет объем производства. В полиплоидном ядре число этих фабрик кратно числу субгеномов. Однако, как правило, фабрики рРНК активны только в одном из субгеномов, а остальные временно неактивны, хотя могут быть активированы в других тканях. Это явление было впервые обнаружено Навашиным в 1934 году21 и с тех пор встречается почти у всех изученных аллополиплоидов.
Клеточное ядро имеет многоуровневую регуляцию. Прежде всего, его размер контролируется десятками генов, определяющих активность различных РНК-полимераз: «Размер имеет значение «22 — Прим. ред.
Пока неясно, по какому принципу отбираются функциональные ядра. Недавно был раскрыт эпигенетический механизм ядерного доминирования, и было показано, что в нем участвуют малые интерферирующие РНК (миРНК), аналогичные тем, которые участвуют в контроле подвижных элементов. Только эта малая РНК не расщепляет мРНК, как у транспозонов, а контролирует метилирование промоторных областей генов рРНК, ингибируя процесс их транскрипции 23 (рис. 4).
Существует термин «аддитивное выражение». Это означает, что эффекты одинаковых генов суммируются при их сочетании. Так, отклонения от этого правила очень часто наблюдаются при аллополиплоидии 24-26. Часто наблюдается изменение паттерна экспрессии (транскрипции) генов, так что в одних тканях гибрида доминирует один ген гомологичной пары, а в других — его двойной ген. Между этими крайностями доминирования существует непрерывный спектр состояний, характеризующихся различным соотношением активности гомологичных генов. Это напоминает чередование времени рабочего дня (онтогенез), когда один ген работает, а его партнер отдыхает в одно и то же время, или оба более или менее задействованы. Логично, не так ли? Некоторые люди более склонны работать в ночную смену, другие — в дневную; задачи, требующие больших усилий (количество белковых продуктов), выполняются более сильным геном, в то время как более слабый ген задействуется при незначительных усилиях.
Аналогия довольно проста и понятна на социальном уровне, но на геномном уровне пока не ясно, почему из двух генов, выполняющих одну и ту же функцию, один работает в одних условиях, а другой — в других. И кто тот невидимый диспетчер, который контролирует их работу? Похоже, что термин «шок» относится не только к геному, но и к его продукту («транскрипционный шок») 20. Это следует из того факта, что лишь небольшая часть генов, претерпевших транскрипционные изменения на ранних стадиях аллополиплоидии, сохранила их в дальнейшем. Большинство генов на более отдаленных стадиях подверглись коррекции своей экспрессии из-за продолжающейся структурной и функциональной дивергенции гомологичных генов.
Структурно функциональная дивергенция генов-гомеологов
Теперь поговорим о дальнейшей судьбе гомологичных генов. Как видно из вышесказанного, оба гомолога в той или иной степени вовлечены в различные стадии онтогенеза аллополиплоидии. Однако, поскольку оба гомолога могут функционировать в диплоидных формах по два (три и т.д.), давление отбора в полиплоиде (по сравнению с диплоидом) меньше, и каждый ген имеет возможность накапливать мутации (рис. 5).
Существует несколько вариантов этого пути 8, 29 (рис. 6). Первая возможность — недостаточная функционализация. В этом случае один гомологичный элемент дополняет другой для более эффективного выполнения конкретной функции в определенных условиях. Это приводит к расширению адаптационного потенциала. Примером может служить ген VRN (ген вернализации) у обыкновенной пшеницы, который определяет восприимчивость к вернализации и время созревания. Каждая из трех гомологичных копий этого гена (соответствующих трем субгеномам A, B и D) имеет ряд аллелей, влияющих на эти физиологические параметры. Сочетание доминантных и рецессивных аллелей на всех трех локусах VRN определяет разнообразие фенотипов с различными сроками вылупления и типами роста (зимние и весенние формы). Это разнообразие внесло значительный вклад в успех обыкновенной пшеницы как всемирной культуры, выращиваемой от умеренных до тропических широт 30.
Тема генетики и селекции сортов культурной пшеницы драматически раскрывается в романе В. Дудинцева «Белые платья». Роман Дудинцева «Белые одежды» описывает конфликт между учеными и руководящими кадрами послевоенного СССР — прим. ред.
Второй путь эволюции гомологичных генов — неофункционализация, при которой один из гомологичных генов приобретает принципиально иную функцию. Этот путь обычно требует более значительных структурных изменений гена, чем первый, и, следовательно, более длительного времени эволюции. Трудно найти пример неофункционализации у полиплоидов, поскольку у синтетических гибридов явно не хватает времени для образования новых генов, тогда как у природных аллополиплоидов — наоборот: долгая эволюция «смяла» и изменила гены настолько, что зачастую довольно трудно определить их происхождение. Наконец, существует путь псевдогеномики, то есть полной потери функции одного из гомологов и превращения его в псевдоген (рис. 6).
Секвенирование всего генома и транскриптома ряда полиплоидов выявило значительную скорость эволюции гомологичных генов и существенные изменения в паттерне экспрессии некоторых из них (неофункционализация), что является косвенным подтверждением указанных выше эволюционных путей 31, 32. У гексаплоидной пшеницы процесс псевдогенерации затронул 38% экспрессируемых генов, что было определено по наличию в них терминирующих кодонов 31.
- 1 Терминология
- 1.1 Типы
- 1.2 Классификация
- 1.2.1 Автополиплоидия
- 1.2.2 Аллополиплоидия
- 1.2.3 Анеуплоид
- 1.2.4 Эндополиплоидия
- 1.2.5 Моноплоид
- 1.3.1 Неополиплоидия
- 1.3.2 Мезополиплоидия
- 1.3.3 Палеополиплоидия
- 1.4.1 Кариотип
- 1.4.2 Гомеологические хромосомы
- 2.1 Животные
- 2.1.1 Люди
- 2.1.2 Рыбы
- 2.2.1 Посевы
- 2.2.1.1 Примеры
Типы
Типы полиплоидии обозначаются в зависимости от количества групп хромосом в ядре клетки. Буква x используется для обозначения количества хромосом в наборе:
- гаплоид (один набор; 1x)
- диплоид (два набора; 2x)
- триплоид ( три набора; 3x), например стерильный шафрановый крокус или арбуз без косточек, также распространенный в филумеTardigrada
- тетраплоид (четыре набора; 4x), например Salmonidae рыба, хлопок Gossypium hirsutum
- пентаплоид (пять наборов; 5x), например Kenai Birch (Betula papyrifera var. Kenaica)
- гексаплоид (шесть наборов; 6x), например пшеница, киви
- гептаплоид или септаплоид ( семь наборов; 7x)
- октаплоид или октоплоид, (восемь наборов; 8x), например, Acipenser (род осетровых рыб), георгины
- декаплоид (десять наборов; 10x), например, некоторые клубника
- додекаплоид (двенадцать наборов; 12x), например, растения Celosia argentea и Spartina anglica или амфибия Xenopus ruwenzoriensis.
Классификация
Автополиплоидия
Автополиплоиды — это полиплоиды с несколькими наборами хромосом, происходящие от одного таксона.
Двумя примерами природных автополиплоидов являются растения Tolmiea menzisii и белый осетр Acipenser transmontanum. В большинстве случаев аутополиплоидия возникает в результате слияния нередуцированных (2n) гамет, что приводит к триплоидному (n + 2n = 3n) или тетраплоидному (2n + 2n = 4n) потомству. Триплоидное потомство обычно стерильно (как в случае эффекта триплоидного блока), но в некоторых случаях оно может производить большую долю нерепродуцирующих гамет, способствуя образованию тетраплоидов. Этот путь к тетраплоидии называется «триплоидным мостом». Триплоиды также могут быть получены путем бесполого размножения. На самом деле, стабильная автотриплексность у растений часто ассоциируется с апомиктическими системами спаривания. В сельскохозяйственных системах автотриплоидия может привести к дефициту семян, как в случае с арбузами и бананами. Триплоидия также используется в разведении лосося и форели для обеспечения стерильности.
В редких случаях автополиплоиды возникают в результате спонтанной дупликации соматического генома, как это наблюдалось у плодоносящих яблонь (Malus domesticus). Это также наиболее распространенный способ искусственного индуцирования полиплоидии, когда такие методы, как слияние протопластов или обработка колхицином, оризалином или ингибиторами митоза, используются для нарушения нормального митотического деления, что приводит к образованию полиплоидных клеток. Эта процедура может быть полезна в селекции растений, особенно при попытке ввести зародышевую плазму на уровне плутидиоза.
Автополиплоиды имеют по крайней мере три гомологичных набора хромосом, что может привести к высокой степени плюрипотентности во время мейоза (особенно у новообразованных автополиплоидов, также называемых неополиплоидами) и относительному снижению фертильности из-за производства анеуплоидных гамет. Естественный или искусственный отбор на плодовитость может быстро стабилизировать мейоз у автополиплоидов путем восстановления димерного спаривания во время мейоза, но высокая степень гомологии между дуплицированными хромосомами приводит к полисомному наследованию у автополиплоидов. Этот признак часто используется в качестве диагностического критерия для отличия аутополиплоидов от аллополиплоидов, которые обычно демонстрируют диомерное наследование после прохождения неопластической стадии. Хотя большинство полиплоидных видов четко характеризуются как автополиплоидные или аллополиплоидные, эти категории представляют собой крайние точки спектра дивергенции между родительскими субгеномами. Полиплоиды, находящиеся между этими двумя крайностями, часто называемые сегментарными аллополиплоидами, могут иметь промежуточные уровни полисомного наследования, которые варьируют в зависимости от локуса.
Кариотипирование
Кариотипирование — это цитогенетический метод, основанный на визуализации хромосом человека с помощью светового микроскопа и выявлении отклонений в их количестве или структуре. Для получения хромосомного препарата лимфоциты культивируют в течение 72 часов в среде, в которую добавлен митоген — вещество, стимулирующее деление хромосом. После культивирования лимфоциты обрабатывают колхицином, который останавливает деление клеток в метафазе. Затем препараты хромосом монтируются на предметные стекла и окрашиваются красителем Giemse для визуализации их структуры и морфологии, а также для выявления дискордантных регионов. Затем подсчитывается количество хромосом и анализируется структура каждой хромосомы 15. В лаборатории Hemotest используется мощная система метафазного исследования Metafer, которая в течение 5 минут проводит поиск 100 метафаз стекла и делает снимки с высоким разрешением, что позволяет получить результаты за очень короткое время. Изображения хранятся в единой базе данных. Metafer предлагает идеальные условия для автоматического и полуавтоматического анализа хромосомных аномалий и способствует объективной и быстрой диагностике генетических дефектов в клетках.
Наряду с преимуществами метод кариотипирования имеет и некоторые недостатки. Во-первых, относительно низкое разрешение микроскопов позволяет выявлять только основные хромосомные аномалии, такие как анеуплоидии, сдвиги Робертсона, сбалансированные сдвиги и инверсии 16. Во-вторых, цитогенетический анализ проводится на живых клеточных культурах 17.
Хромосомный микроматричный анализ
Анализ хромосомных микрочипов (CMA) — это технология молекулярно-генетического скрининга, позволяющая с высокой точностью определить количество и тип хромосомных аномалий и выявить хромосомные аномалии длиной от 50 до 100 тысяч пар нуклеотидов, такие как микроделеции и микродупликации, что в десять раз больше, чем при анализе кариотипа18. Поэтому преимуществами СМА являются высокое разрешение, более короткий цикл исследования, более объективные результаты и отсутствие необходимости в культуре клеток.
Одним из преимуществ СМА перед классическими цитогенетическими методами диагностики хромосомных аномалий является способность обнаруживать и идентифицировать хромосомный дисбаланс с высокой точностью. Это важно не только для субмикроскопических аномалий, но и для видимых аномалий неясного происхождения, таких как маркерные хромосомы и несбалансированные перестройки. Для этого в лаборатории «Гемотест» используются ДНК-микрочипы и анализатор Genoscan 3000, одобренный в России.
Несмотря на ряд преимуществ, этот метод имеет и свои недостатки. В СМА, например, ДНК пациента анализируется в сравнении с нормальной ДНК, поэтому дисбаланс может быть обнаружен только в сравнении с контролем, в то время как сбалансированные перестройки не могут быть обнаружены. Наличие сбалансированной перестройки у одного из партнеров не имеет клинического значения, но является предиктором повышенного риска выкидыша при последующих беременностях, если один из родителей является носителем.
Клиническое наблюдение
В семье были частые выкидыши. В семье было 3 выкидыша в первом триместре. Семейная история спонтанных абортов была определена путем сбора семейных данных. Кариотипирование в лаборатории «Гемотест» выявило сбалансированный сдвиг в хромосомах 16 и 19, унаследованных от матери, что может привести к хромосомным аномалиям у плода. Учитывая возраст женщины (35 лет), ее историю болезни и высокий риск повторных спонтанных абортов, паре было рекомендовано ЭКО с предимплантационной диагностикой хромосомных аномалий у эмбриона перед переносом. Были отобраны эмбрионы без хромосомных аномалий, и перенос прошел успешно. Беременность завершилась рождением здоровых близнецов.
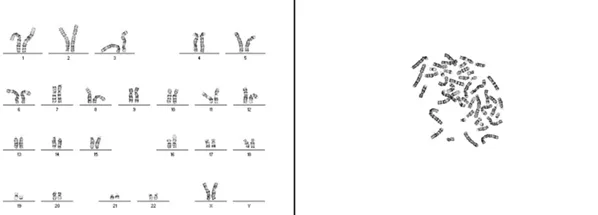
Рисунок 2. Кариотип пациента N — 46,XX, t(16;19)(p13.3;q12)
Полиплоидия у растений
Наличие более одного генома в одном ядре сыграло важную роль в происхождении и эволюции растений и, возможно, является самым важным цитогенетическим изменением в разнообразии и эволюции растений. Растения стали воротами к познанию клеток с более чем двумя наборами хромосом на клетку.
С первых дней подсчета хромосом было обнаружено, что большое количество диких и культурных растений (включая некоторые из наиболее важных) являются полиплоидными. Почти половина известных видов искусственных растений (цветковых) являются полиплоидными, как и большинство папоротников (95%) и большое количество мхов.
Наличие полиплоидии у гимноспермов встречается редко и сильно варьирует между группами наземных растений. В целом, полиплоидные растения отличаются высокой адаптивностью, они способны занимать места обитания, которые не могли занять их диплоидные предки. Кроме того, полиплоидные растения накапливают больше «вариаций» за счет большого количества копий генома.
В растениях аллополиплоиды (которые более распространены в природе), возможно, сыграли фундаментальную роль в диверсификации и адаптивной радиации во многих группах.
Улучшение садоводства
У растений полиплоидия может возникать в результате ряда явлений, наиболее распространенным из которых, пожалуй, являются ошибки в процессе мейоза, приводящие к образованию диплоидных гамет.
Более 40% культурных растений являются полиплоидными, например, люцерна, хлопок, картофель, кофе, клубника и пшеница, при этом между одомашниванием растений и полиплоидией нет никакой связи.
С тех пор как колхицин был использован в качестве агента, индуцирующего полиплоидию, он применялся в сельскохозяйственных культурах в основном по трем причинам:
-вызвать полиплоидию у некоторых важных видов для получения лучших растений, поскольку полиплоиды обычно имеют фенотип, в котором количество «гигабайтов» значительно увеличивается из-за большего числа клеток. Это привело к значительным достижениям в садоводстве и генетическом улучшении растений.
-Гибриды полиплоидизируют и восстанавливают фертильность для рециклирования или синтеза определенных видов.
-и, наконец, как средство передачи генов между видами с разным богатством или внутри одного вида.
Примеры в растениях
Естественным полиплоидом, имеющим большое значение и особый интерес среди растений, является обыкновенная пшеница, Triticum aestibum (гексаплоид). Кроме ржи, был специально создан полиплоид под названием тритикале, аллополиплоид с высокой урожайностью пшеницы и высокой урожайностью ржи с большим потенциалом.
Пшеница играет чрезвычайно важную роль в зерновых культурах. Существует 14 видов пшеницы, которые эволюционировали путем аллополиплоидии, образуя три группы: с 14, 28 и 42 хромосомами. Первая группа включает самые старые виды рода. T. monococcum Y T. boeoticum.
Вторая группа состоит из 7 видов и, по-видимому, возникла в результате гибридизации T. boeoticum с диким видом травы другого рода под названием Egilops. В результате скрещивания получается высокостерильный гибрид, который может привести к фертильному аллотетраплоиду путем дупликации хромосом.
Третья группа из 42 хромосом — это хлебная пшеница, которая, вероятно, появилась в результате скрещивания триплоидного вида с другим видом Egilops и последующего удвоения общего числа хромосом.







