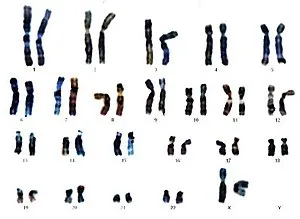
Каждый человек имеет 46 хромосом, состоящих из 23 пар. Двадцать две пары почти идентичны, но в последней паре есть существенные различия. Это хромосомы, которые регулируют половую принадлежность человека.
Хромосома

Форма строения постнатальных хромосом — средняя стадия митоза. 1, хроматиды — 2, центромеры — 3, короткие плечи — 4, длинные плечи.
Хромосомы (греч. «тело» и «тело») — это ядерные белковые структуры в ядре эукариотических клеток, которые становятся хорошо видимыми на определенных стадиях клеточного цикла (митоз или редукция). Хромосомы представляют собой высокую концентрацию хроматина, постоянно присутствующего в клеточном ядре. Изначально этот термин был придуман для обозначения структур, обнаруженных в эукариотических клетках, но в последние десятилетия его стали относить к бактериальным хромосомам. Хромосомы содержат большую часть наследственной информации.
Содержание
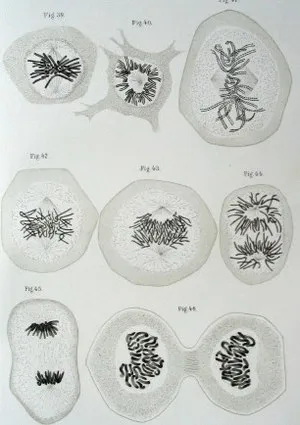
Фотография из книги В. Флеминга, показывающая различные стадии деления эпителиальных клеток у саламандр (W. Fleming, Zellsubstanz, Kern und Zelltheilung, 1882).
В различных статьях и книгах приоритет открытия хромосом отдается разным людям, но в большинстве случаев в качестве года открытия хромосом называется 1882 год, а в качестве первооткрывателя упоминается немецкий анатом В. Флеминг. Однако не будет преувеличением сказать, что не он открыл хромосомы, а в своей основной книге «Zellsubstanz, Kern und Zelltheilung» (на немецком языке) он собрал и систематизировал информацию о хромосомах и дополнил ее результатами собственных исследований. Термин «хромосома» был введен немецким блоггером Генрихом Вальдмейером в 1888 году. Хромосома» буквально означает «цветное тело», так как основные пигменты хорошо связаны с хромосомами.
Сегодня трудно сказать, кто первым описал и сконструировал хромосомы. В 1872 году швейцарский ботаник Карл фон Нагили опубликовал работу, в которой показал, что тела некоторых видов появляются в ядерном положении во время деления клеток при образовании пыльцы у лилии (Lilium tigrinum) и рода Мурасаки-цуцую (Tradescantia). Однако его конструкция не позволяет однозначно утверждать, что хромосомы являются теми, которые видел К. Нагири. В том же 1872 году ботаник Э. Руссоу опубликовал план деления клеток при образовании спор в пыльце папоротников и лилий рода Ophioglossum (Liliumbulbiferum). На их иллюстрациях можно легко идентифицировать отдельные хромосомы и стадии деления. Однако некоторые исследователи считают, что немецкий ботаник Вильгельм Гофмейстер впервые увидел хромосомы в 1848-1849 годах, задолго до К. Нагили и Э. Руссоу. Однако ни К. Нагили, ни Э. Руссоу, ни тем более В. Хоффмайстер не осознали важности увиденного.
После повторного открытия законов Менделя в 1900 году потребовался всего год или два, чтобы показать, что хромосомы работают так, как и ожидалось от «наследственных частиц». Т.Бовери и Уолтер Саттон (1902-1903) первыми независимо друг от друга выдвинули гипотезу о генетической роли хромосом в 1902 году. полный набор хромосом. Он также обнаружил, что состав разных хромосом не одинаков. У. Сеттон изучал гаметогенез у саранчи Brachystolamagna и понял, что поведение хромосом во время мейоза и оплодотворения полностью объясняет картину отклонений от менделевских законов и образования новых комбинаций.
Экспериментальное подтверждение этих идей и окончательная формулировка хромосомной теории были сделаны в первой четверти 20 века основателями классической генетики, работавшими в США с Drosophila melanogaster (D. melanogaster): T. Morgan, C.B. Bridges, A.-H. Стертевант и Х. Моллер. На основе своих данных они сформулировали «хромосомную теорию наследственности». Согласно ей, передача генетической информации связана с хромосомами, где гены расположены в линейных последовательностях. Эти результаты были опубликованы в 1915 году в книге «Механизмы законов Менделя».
В 1933 году Т. Морган был удостоен Нобелевской премии по физиологии или медицине за открытие роли хромосом в наследственности1.
Хромосомы эукариот
В основе хромосом лежат линейные (ациклические) макромолекулы дезоксирибонуклеиновой кислоты (ДНК) значительной длины (например, молекулы хромосомной ДНК человека содержат от 50 до 245 миллионов пар азотистых оснований). В растянутом состоянии хромосомы человека могут достигать 5 см в длину. Хромосомы также состоят из пяти специфических белков (H1, H2A, H2B, H3 и H4 (называемых гистонами)) и нескольких негистоновых белков. Аминокислотная последовательность гистонов высоко консервативна и мало отличается между различными группами организмов.
Во время интерфазы хроматин не обогащается, но его клон по-прежнему представляет собой ДНК-белковый комплекс. Макромолекула ДНК оборачивается вокруг октамера (структуры, состоящей из восьми белковых шариков) из гистоновых белков H2A, H2B, H3 и H4, образуя структуру, называемую нуклеосомой. В целом, вся конструкция немного напоминает бусы. Последовательности таких нуклеосом, которые связываются с белком H1, называются нуклеозидными нуклеотидами или филаментами и имеют диаметр около 10 нм.
В ранней средней фазе (G1), каждая будущая хромосома основана на одной молекуле ДНК. В фазе синтеза (S) молекула ДНК вступает в процесс репликации и реплицируется. В поздней промежуточной фазе (фаза G) позвоночник каждой хромосомы2Основа каждой хромосомы формируется в результате репликации и состоит из двух идентичных молекул ДНК, соединенных вблизи центромеры.
Перед началом деления клетки хромосомы, теперь представленные нитями нуклеосом, начинают закручиваться в спираль или накапливаться и формировать более толстые нити хроматина (хроматин) с помощью белка H1. 30 нм. в результате дальнейшей спиральной конфигурации, в течение среднего периода диаметр хромоида 700 нм. Значительная толщина (диаметр 1400 нм) хромосом на средней стадии в конечном итоге становится видимой под световым микроскопом. Две хромосомы, получившиеся в результате репликации, по-прежнему тесно связаны друг с другом, поэтому конденсированные хромосомы имеют Х-образную форму (часто с неровными плечами) (более подробную информацию о судьбе хромосом во время деления клетки см. в статье о митозе и редукции).
Первичная перетяжка
Хромосомный вклад (Х.п.), где расположена центромера, разделяющая хромосомы на плечи.
Вторичные перетяжки
Морфологические признаки, позволяющие идентифицировать отдельные хромосомы внутри набора. От первичных перестроек их отличает отсутствие выраженных углов между сегментами хромосом. Вторичные перестройки бывают короткими и длинными и располагаются в разных точках хромосомы. У человека это хромосомы 9, 13, 14, 15, 21 и 22.
Типы строения хромосом
Существует четыре типа структуры хромосом:.
- телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
- акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
- субметацентрические (с плечами неравной длины, напоминающие по форме букву L);
- метацентрические (V-образные хромосомы, обладающие плечами равной длины).
Тип хромосомы постоянен для каждой сцепленной хромосомы и может быть стабильным у всех представителей одного вида или рода.
Гены, наследуемые отцом и матерью, вносят 50% и 50% в хромосомный состав потомства. В каждой клетке тела человека имеется 23 пары хромосом. Другими словами, каждая клетка имеет 46 хромосом.
Четыре возраста Y-хромосомы
Первый возраст. Дата рождения. Как уже говорилось, пол определяется не всеми хромосомами животных. Расовые хромосомы возникают, когда в результате мутации пол определяется одним геном, а не набором многих генов. Важно отметить, что этот ген не отвечает исключительно за все гендерные характеристики. Многие гены должны работать вместе, чтобы обеспечить полноценное развитие как мужского, так и женского организма. Большинство из них находятся на аутосомах. Продукт половых генов просто действует как спусковой крючок и определяет ход развития того или другого. Если доминантный аллель вызывает развитие мужского пола, то хромосома, на которой находится половая хромосома (и ее X-аналог), окрашивается. Если этот новый доминантный аллель направляет рост по женскому пути, хромосома переносит его в хромосому W (и ее аналог в Z).
В эволюции млекопитающих это произошло один раз у общего предка марсиан и плаценты, около 180 миллионов лет назад. Таким образом, Y-хромосома всех млекопитающих (от кенгуру до человека) происходит от одной аутосомы, в которой один из аллелей гена SOX3 в результате мутации стал мужским основным мембранным (обращенным к полу Y) геном. Примерно в то же время или позже (около 140 миллионов лет назад) птицы приобрели систему полового детерминирования ZW. Она также основана на одной аутосомной хромосоме, раз
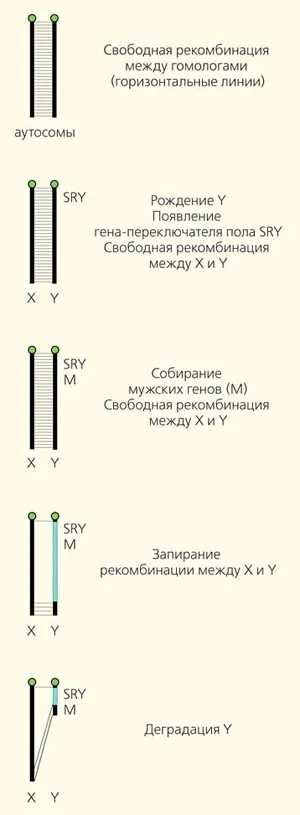
Рисунок 2: Эволюция Y-хромосомы
Рыба — это совсем другое дело. У рыб племенные хромосомы (как Y, так и W) часто возникают в разное время и основываются на совершенно разных аутосомах. В большинстве случаев все это произошло относительно недавно. Системы определения пола различаются у разных видов, даже в пределах относительно небольшого семейства рыб Pisilia. Например, у гиппий — система XY, а у черных моллюсков — система ZW, которые имели общего предка всего 20 миллионов лет назад. Ни в каком виде! У мечей есть популяции с XY и ZW системами полового детерминирования и нехромосомными системами полового детерминирования в пределах одного вида.
Почему это так; почему половые хромосомы рыб намного моложе, чем половые хромосомы птиц? Самое простое объяснение заключается в том, что они намного больше наших. Те, кто имел расовые хромосомы, давно уже исчезли (есть более духовный случай под названием «источник молодости», о котором мы поговорим чуть позже).
Как справедливо заметил Козьма Прутков, «первый шаг младенца — это первый шаг к его смерти». Как только гипертензия становится Y-хромосомой, неумолимая логика физического отбора приводит к медленному и неизбежному процессу распада (Рисунок 2). Каждый шаг эволюции приводит к повышению приспособленности, по крайней мере, к сохранению приспособленности предка, и в результате этих шагов вся хромосома деградирует.
Вторичный период. Детский и мужской набор генов. Гены, играющие важную роль в жизни мужчины, начинают накапливаться на Y-хромосоме. Например, у человека, помимо определяющего пол гена SRY, существуют гены, участвующие в сперматогенезе и формировании семенников. У гуппи Y-хромосома содержит гены, участвующие в яркой окраске тела и привлекательной форме плавников, которые являются элементами брачного наряда (рис. 3 и 4).
Y-хромосоме столько лет, на сколько она выглядит
У рыбок гуппи Y-хромосома была обнаружена в 1934 году. Ученые заметили, что некоторые цветовые особенности передавались только в отцовской популяции и никогда не проявлялись у самок. Однако в то время методы подготовки хромосом не позволяли идентифицировать половые хромосомы. Они были очень похожи друг на друга. Только в 1990 году, используя специальное пятно, исследователи смогли различить X и Y хромосомы Габи. Было показано, что Y несет большие блоки плотно упакованной ДНК (гетерохроматина) в удаленных (т.е. вдали от центриолей) регионах, которые отсутствуют в Х-хромосоме (которая немного длиннее своего Y-аналога). Авторы предположили, что нерекомбинантная Y-специфическая часть гуппи, содержащая мужской ген, расположена именно в этом удаленном регионе6.
Впоследствии, используя флуоресцентную гибридизацию in situ (FISH), они обнаружили, что эта область содержит последовательности ДНК, специфичные для Y-хромосомы. Окрасив мужские и женские гаппи различными флуоресцентными красителями, исследователи нанесли ДНК на образцы хромосом. Благодаря сродству между идентичными последовательностями ДНК, окрашенные образцы связываются с хромосомной ДНК. Дистальная область Y-хромосомы светилась одним цветом, в то время как остальная часть генома светилась обоими цветами. Это означает, что дистальная часть содержала уникальные последовательности ДНК, которых не было в женском геноме. Это подтвердило вывод о том, что трибиальные хромосомы Gap разделены на две части: гомологичную проксимальную часть (около центрального тела) и особую отдаленную часть7.
Таким образом, Y-хромосома гуппи жива уже в первых двух возрастах. Это означает, что мужские гены были рождены и собраны. Когда она вступила в пожилой возраст, произошло ли у нее торможение рекомбинации между этими генами и генами, определяющими пол? Если да, то насколько велика зона табу?
Исследования редуцированных хромосом могут ответить на этот вопрос. В 1995 году один из нас (и его коллеги) провел электронно-микроскопический анализ брачных половых хромосом самцов гуппи и определил (независимо друг от друга), что X и Y полностью гомологичны друг другу. Это казалось весьма логичным. В большинстве из сотен проанализированных клеток все хромосомы были одинаковыми по всей своей длине. Несколько пар хромосом с кажущимися неполными синапсами и слегка отличающейся длиной хромосом были расценены одним из нас как ложные артефакты8.
Из результатов исследования электронной микроскопии, опубликованных в 2001 году. По мере продвижения выводов самая длинная хромосома Y вынуждена уменьшаться, пока хромосомы не сравняются. Авторы предположили, что спаривание начинается на близлежащем скрепленном участке, а затем распространяется на удаленный нескрепленный участок. Поскольку используемый метод не позволяет отличить проксимальный конец от дальнего, авторы определили, что эти крайние синапсы7 одинаковы, если концы x и y различны, а проксимальный конец одинаков.
Однако этому выводу противоречат наблюдения того же автора за хромосомами самца гуппи для метафоры I (см. рис. 1). На этой стадии племенные хромосомы всегда сцеплены с удаленным концом, чего не должно происходить, если в удаленном участке нет рекомбинации. Точки соединения также наблюдались проксимально, но очень редко. Чтобы примирить края, авторы предположили, что спаривание отдаленных конечностей происходит не за счет рекомбинации, а за счет другого механизма.
Реабилитация юной Y-хромосомы
Чтобы устранить эти несоответствия, они решили изучить расовые хромосомы гуппи, используя мужские эмбрионы с иммунитетом к основному белку (этот метод не использовался в предыдущих исследованиях)****. Они использовали препараты тестикул гуппи и антитела, помеченные флуоресцентными красителями для двух белков — SYCP3 и MLH1. Первый белок формирует хромосомный вал, а второй маркирует точку рекомбинации. ДНК была покрыта синим флуоресцентным красителем DAPI. В результате была отмечена точка рекомбинации и получено изображение билатеральных хромосом (парных хромосом) (рис. 5, А). В каждой ячейке было 23 билатерали, каждая из которых имела по крайней мере одну точку рекомбинации. Какие из них образованы X и Y хромосомами и где находятся проксимальные и дистальные концы билатеральных соков, т.е. центры составляющих их хромосом?
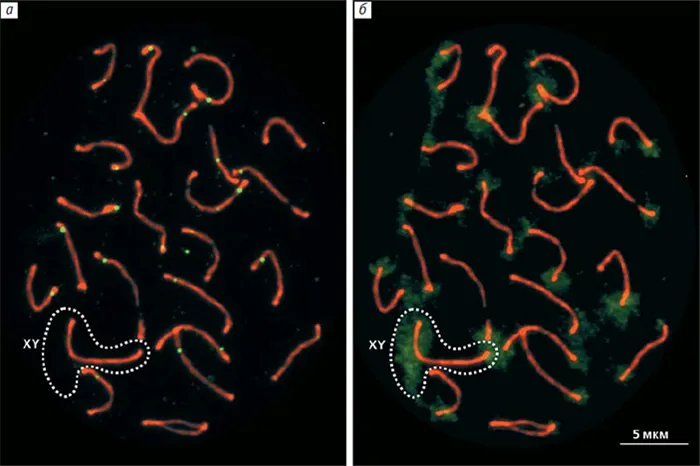
РИСУНОК 5. Дублирующие хромосомы у самцов хлеба, полученные иммуноабузусом двух белков SYCP3 в хромосомах, маркирующих точку рекомбинации, и в редукции, формирующей вал MLH1. Используя антитела, помеченные флуорохромами (красный для белка вала и зеленый для индекса MLH1), можно было определить положение этих точек между всеми двумя странами (a). В тех же клетках красного дерева после реакции рыбы рядом с каждой центрифугой были обнаружены двумерные области гетерохроматина (диффузный зеленый сигнал). Место укуса XY также определялось с помощью рыбы. На удаленном конце располагалась большая область гетерохроматина 5.
Флуоресцентная гибридизация in situ ДНК самцов гуппи с хромосомами красного дерева была использована для идентификации двустороннего XY. Образцы ДНК, помеченные зеленым флуорохроматическим цветом, наносились на препараты хромосом, и метод использовался для выявления гетерохроматиновых областей хромосом, особенно тех, которые содержат компактную ДНК. Это блок контрамерного гетерохроматина во всех хромосомах, включая X и Y хромосомы, и очень большой блок гетерохроматина на удаленном конце двойного пола (рис. 6, B). Во время экспансии половых клеток при неполных концентрациях помеченные районы спаривались, а проксимальные концы хромосом X и Y оставались свободными (рис. 6, A и B). Так выглядело большинство билатеральных зародышевых клеток в неполной концентрации.
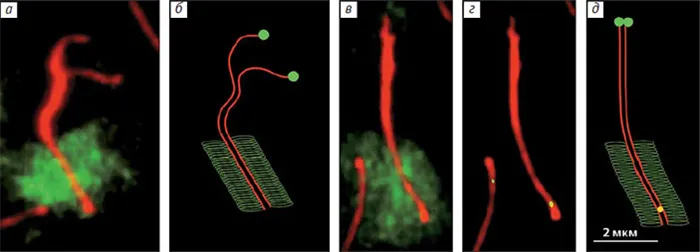

РИСУНОК 5. Дублирующие хромосомы у самцов хлеба, полученные иммуноабузусом двух белков SYCP3 в хромосомах, маркирующих точку рекомбинации, и в редукции, формирующей вал MLH1. Используя антитела, помеченные флуорохромами (красный для белка вала и зеленый для индекса MLH1), можно было определить положение этих точек между всеми двумя странами (a). В тех же клетках красного дерева после реакции рыбы рядом с каждой центрифугой были обнаружены двумерные области гетерохроматина (диффузный зеленый сигнал). Место укуса XY также определялось с помощью рыбы. На удаленном конце располагалась большая область гетерохроматина 5.







